热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
讨论
场通量
在该领域,N2Ofluxes 平均为 6.7 6 2 lmol.m-2.h-1,这在其他潮汐研究使用更常见的室通量方法报告的通量范围内(Middelburg 等人 1995,Wang 等人 2007,Adams et al.2012)。然而,我们的范围大于潮汐平板文献中报道的范围(表 1)。由于方法上的差异,这可能并不令人惊讶。室通量法测量在更大区域内整合的气体通量(通常为 0.1 m2;Middelburg et al.1995,Wang et al.2007),而微剖面方法基于仅在沉积物中的一个点采集的剖面对气体通量进行建模。因此,来自微剖面方法的通量被整合到一个小得多的区域,导致更多的变量值。出乎意料的是,几乎所有的通量都是负的,暴露的沉积物越长,吸收越多(图 1)。这尤其令人惊讶,因为脉冲事件,特别是润湿/干燥动力学,通常会导致来自陆地的 N2O 流出量增加 油(Kim 等人,2012 年)。然而,我们调查的外部潮汐脉冲是这些潮滩沉积物中养分可用性的主要驱动因素。因此,我们假设退潮代表了这些沉积物营养物质快速剥夺的时期。
最初,在检查脉冲潮汐暴露对潮间带 N2Ofluxes 的影响时,我们预计整个低潮期间沉积物中的氧气动力学会发生变化。特别是,我们预测,沉积物暴露在大气中的时间越长,氧气渗透到沉积物中的深度就越深.反过来,这种增加的氧气供应可能会刺激硝化作用,从而增加低潮期间 N2O 的产量和随后的排放量。相反,我们观察到沉积物氧气渗透深度随暴露时间没有显着变化(表 1)。这可能是一个结果沉积物含水量(SWC),在整个低潮期间几乎保持不变,并且总是处于或非常接近饱和状态(表 1)。这表明,尽管沉积物暴露在空气中,但沉积物孔隙空间仍然充满水,并不显着重新充氧,表明低潮时的 N2Oflux 不是由硝化器的氧气供应变化驱动的,正如我们最初怀疑的那样(表 1,图 1)。
此外,所有产生 N2O 的过程都依赖于向沉积物供应溶解的无机氮,因此,如果 DIN 浓度耗尽,这些过程的速率会减慢,N2O 的产生也会减慢(Burgin 和 Hamilton 2007)。然而,反硝化可以适应通过吸收 N2O 并将其用作终端电子受体来降低底物可用性(Miller 等人,1986 年)。因此,我们假设 N2O 吸收随沉积物暴露时间显着增加是由反硝化作用驱动的退潮时微生物群落经历的营养缺乏(图 1)。这种 N2O 消耗的增加代表了该系统中脉冲的显着影响。作为这种营养驱动脉冲的证据,我们可以看看 N2Oflux 和沉积物需氧量。7 月 31 日 N2Oflux 与 SOD 的关系与 8 月 8 日不同;两者均显着关系(P,0.05),但对于相同的 SOD, 8 月 8 日的 N2O 吸收量比 7 月 31 日多(P = 0.05)。我们认为这两个样本日期表现出对比鲜明的沉积物 N2O 动态,因为它们经历了非常不同的天气条件,并且这种天气变化导致了内部竞争动态的转变。沉积物。7 月 31 日多雾,有微量降水(NOAA,数据可在线获取),5 平均沉积物温度为 24.28C,而 8 月 8 日炎热晴朗,平均沉积物温度为 30.58C。这两天没有显着差异,因此,N2Oflux 的差异可能不仅仅是温度变化的结果。我们假设在阳光明媚的日子里 N2O 吸收增加是由于反硝化器必须与微藻群落竞争用于养分的泥滩;即低潮时的硝酸盐。这些沉积物中含有丰富的沉积物微藻群落;它们是以光合硅藻为主的组合(称为微底栖植物 [MPB]),占据了 t 潮滩上只有几毫米的沉积物(Pascal 等人,2013 年)。作为光合自养生物,MPB 需要包括氮在内的无机养分才能发挥作用,因此不断与细菌(包括反硝化菌)竞争沉积物中的养分。因此,我们建议多云时,如 7 月 31 日,由于光合速率降低,MPB 养分需求相对较低。这将为包括反硝化菌(产生 N2 或 N2O)在内的细菌留下更多孔隙水养分。然而,在晴天,由于光合作用速率高,反硝化器必须与 MPB 竞争硝酸盐(Sundback 等人,2000 年)。在这些条件下,孔隙水硝酸盐可能会更快耗尽,尤其是在退潮时,驱使反硝化器更加依赖 N2O 作为其终端电子受体(Miller et al.1986)。这进一步表明,随着沉积物潮汐暴露的时间越长,养分的可用性和竞争可能会推动 N2O 的吸收增加,就像 鉴于其消耗 N2O 的能力,N2O 吸收的增加是潮滩沉积物提供的生态系统服务,并且是潮汐淹没的日常脉冲事件,随后暴露在大气中对潮滩氮动力学的显着影响。
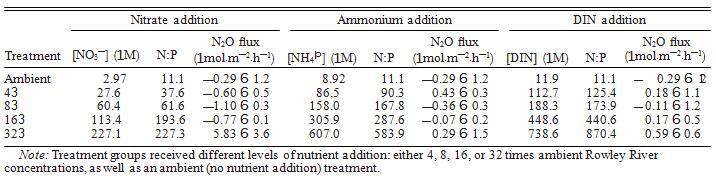
表 2. 每种处理的养分处理浓度、溶解无机氮 (DIN) 与溶解无机磷 (DIN DIP) 的比率,以及每个实验处理和处理水平下淹没核心的 N2O 通量。
营养添加实验
为了区分我们在野外观察到的 N2Oflux 的特定养分驱动因素,我们在相同潮间带沉积物的额外核心上设计了一个补充养分添加实验。这些添加的 N2Ofluxes 的量级通常比我们观察到的要低得多场(P,0.01)。这可能是沉积物整体代谢率下降的结果,因为 7 月和 8 月的平均沉积物温度高于 9 月(分别为 27.58C 和 178C)。温度下降也导致与现场测量的核心相比,养分添加核心中的 SOD 显着降低(P = 0.01)。因此,我们观察到的养分添加响应的总体趋势可能是夏季温度下发生的情况的保守近似值。
总体而言,我们发现当核心被处理淹没时 N2Ofluxs 显着高于核心暴露在空气中时(P,0.01;图 2)。这与我们的实地观察一致,并支持我们的结论,即 N2O 吸收随着暴露而增加竞争耗尽的养分供应。然而,无论处理如何,对裸露的岩心都没有显着的处理效果(图 2b),表明处理效果是暂时的,只有在沉积物被淹没时才会持续。一旦暴露,孔隙水养分似乎快速使用,驱使反硝化器越来越依赖间隙和大气 N2O 进行反硝化,从而导致 N2O 净负通量进入沉积物(图 2b)。
3. (a) DIN (y 0.0009x 0.18, R2 0.82, P 0.034) 和 (b) DIN DIP 添加量 (y 0.072x 1.5, R2 0.71, P 0.07) 的淹没 N2O 通量与 N 与 P 的比率。 箭头表示处理方向(从低到高的营养添加)。 误差线表示 6SE,请注意不同的比例。
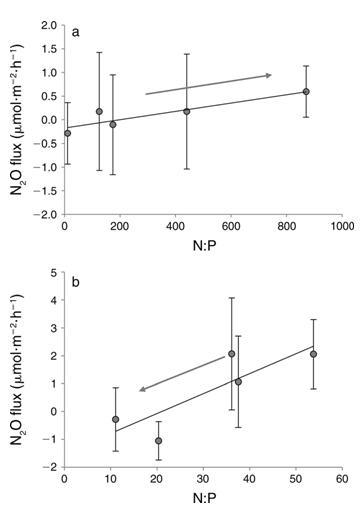
表 2. 扩展。
当核心被处理淹没时,只有硝酸盐和 DIN 处理在所有核心中都有显着的处理效果,尽管 DINþDIP 在所有核心中都是显着的(图 2a)。铵处理没有显示出处理效果,N2Oflux 与 N2Oflux 之间没有显着关系和铵浓度(表 2)。这可能是由于 Michaelis-Menten 动力学。N2O 可以在硝化的第一步产生,氨氧化,这个过程可以通过氨氧化细菌 (AOB) 或氨进行氧化古细菌(AOA)。培养物中的AOA(Candidatus Nitrosopumilus maritimus'strain SCM1)表现出非常低的饱和值(Km);130 nmol/L NH4þ,而AOB具有更高的值(Km'100–550 lmol/L NH4þ; Martens-Habbena et al.2009。鉴于这些沉积物即使对非常高的铵添加量(超过 600 lmol/L)也没有表现出任何反应,我们假设这些沉积物中的硝化群落以 AOA 为主,正如已经显示在一些农业土壤中(Leininger et al.2006。因此,我们认为这些沉积物中的硝化器在非常低的铵浓度下饱和,使它们无法使用额外的底物来提高硝化速率,因此 N2O 的产生是有限的。硝化器缺乏响应添加铵盐也表明硝化作用不是调节这些沉积物中 N2O 通量的过程,而且信号更可能由反硝化作用和/或 DNRA 支配,如硝酸盐和 DIN 添加物的显着处理效果所示(图.2). 已发现反硝化作用是温带(例如,Davidson 等人 1986)和热带森林土壤(例如,Bowden 等人,1992 年,Silver 等人,1999 年)以及在一些海洋生态系统中(例如,Dong et al.2002,Wang et al.2007)。
硝酸盐处理与 N2Oflux 和硝酸盐浓度之间存在抛物线关系。然而,这种关系完全由 323 处理的 N2Oflux 驱动(5.83 6 3.6 lmol.m—2.h—1),从 0 到 163 的相同关系不是显着表明沉积物群落对硝酸盐添加没有反应,直到它们比环境水平大得多。事实上,除了 323 处理外,所有硝酸盐添加处理都表现出 N2O 吸收(图 2a,表 2)。反硝化和 DRNA可以吸收硝酸盐并产生N2O,尽管每个反应的动力学不同。反硝化在;5至10 lmol/L NO3—饱和(具有Km值),而DNRA在;100至500 lmol/L NO3饱和—(Jorgensen 1989)。鉴于这些 Km 值,我们认为是 DNRA 而不是反硝化,通过产生 N2O 对高水平的硝酸盐做出反应。这与表明 DNRA 可能在附近盐沼中胜过反硝化的研究结果一致潮汐小溪 用硝酸盐施肥(Vieillard 和 Fulweiler 2012)。 从系统中去除哪些生物可利用的氮(例如,Kieskamp et al.1991)。相反,DNRA 的最终产物是铵,氮的另一种生物可利用形式。因此,反硝化有可能抵消增加的 N 负荷,而 DNRA 保持反应性N 在系统内循环。然而,由于动力学,DNRA 可能会在较高的 NO3 浓度下胜过反硝化(Jorgensen 1989)。这表明增加 N 负荷有可能诱导沉积物群落从反硝化向 DNRA 转变,导致进一步的DIN 水平升高时,生态系统中生物可利用 N 的增加以及沉积物 N2O 排放量可能增加。由于这两个过程经常在沉积物中同时发生,因此通常很难区分它们对 N2O 通量的单独贡献。此外,DNRA 是很少专门测量 N2O 排放;然而,有强有力的证据表明 DNRA 在陆地土壤中的 N2O 排放中起着重要作用,尤其是那些高 pH 值的土壤 (. 6.5) 和高碳含量 (C:N.4;Stevens et al.1998,Morley and Baggs 2010)。
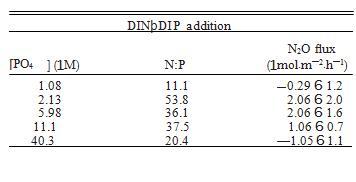
添加 DIN 也有显着的处理效果,N2Oflux 和 DIN 浓度之间呈正线性相关,表明随着水柱 DIN 的增加,沉积物 N2O 的产生也增加(图 2a,表 2)。这遵循类似的线性模式,具有以前在潮间带沉积物中看到过(Middelburg et al.1995),而不是在河口沉积物中看到的指数关系(Seitzinger and Nixon 1985)。这些发现一起表明沿海生态系统的这两个组成部分可能对养分负荷有不同的反应。河口沉积物对养分富集的指数响应(Seitzinger 和 Nixon 1985)表明河口 N2O 通量可能比潮间带沉积物对养分负荷的变化更敏感。已经认为河口沉积物几乎是环境中 N2O 的唯一来源(Middelburg 等人.1995 年),而对人为 N 的更高敏感性可能只会加剧这种来源(Kroeze 等人,2005 年)。 本研究和其他人(例如,Wang 等人,2007 年)已经表明,dalflats 能够成为 N2O 的重要汇。由于对 DIN 负载的线性响应,tidalflat N2Ofluxes 可能对 N 的人为增加不太敏感负载比河口高,但增加的 N 负载仍然有可能将这些系统转移到更持久的 N2O 来源。 2). 令人惊讶的是,这种趋势与单独的 DIN 处理相反。我们发现,随着磷酸盐浓度的增加,更多的 N2O 被吸收(图 2a)。这种关系表明,这些沉积物中 N2O 吸收的过程是 P 限制的。 P 限制的进一步证据是 DIN 和 DINþDIP 处理中 N2Oflux 和 N:P 之间的关系(图 3)。两种处理都显示 N2Oflux 和 N:P 之间的正线性相关(图 3)。然而,斜坡 这些关系的不同之处在于,更重要的是,处理进程对 N2Oflux 具有相反的影响(图 3)。仅添加 DIN,导致 N:P 增加,产生更多 N2O(图 3)。 3a). 相反,在 DINþDIP 添加中,DIP 的添加和 N:P 的同时减少,吸收的 N2O 越多(图 3b)。这些数据一起表明这些沉积物中的 N2O 吸收可能是磷 -劳斯有限。
虽然氮通常被认为是海洋生态系统中的限制性营养物质,但在整个海洋环境中的各种群落中都发现了磷限制(Fourqurean 等人 1992、MacRae 等人 1994、Cotner 等人 1997、Karl 和 Yanagi 1997,Moore et al.2013),并且在来自同一生态系统的潮汐溪沉积物中也发现了 P 限制的证据(Vieillard 和 Fulweiler 2012)。此外,已经发现盐沼沉积物的异养微生物群落主要受到限制由磷(Sundareshwar 等人,2003 年)。Sundareshwar 等人(2003 年)还表明,当磷限制时 N2O 生成率升高,但在添加磷时降低,这表明反硝化器的限制,正如我们在本研究中观察到的(图 3;Sundareshwar 等人 2003。我们的结果表明,这些潮滩沉积物中反硝化器对 N2O 的吸收也可能受到 P 限制。在我们的营养修正处理中,硝酸盐的存在或不存在,理论上 反硝化 N2O 吸收的驱动因素(Miller 等人,1986 年),不会显着影响 N2O 吸收。吸收率只会在 P 浓度增加时急剧增加。这表明反硝化和 DIP 之间存在基本且相对未开发的联系。它还表明与单纯的硝酸盐浓度相比,N:P 的比例对 N2O 吸收的影响可能更大。

图 1. Rowley River 现场采样点,2012 年 6 月 29 日。照片来源:AM Vieillard。
该研究表明,较低的 N:P 比率有利于潮滩沉积物中 N2O 的吸收,而较高的 N:P 比率会刺激 N2O 的净产量(图 3)。这一发现对沿海生态系统具有潜在的深远影响。虽然人类活动如肥料生产影响氮和磷循环,在全球范围内,N 的自然和人为来源比 P 对环境大得多(例如,Fanning 1989,Carpenter 和 Romans 1991,Karl 等人,1997)。因此,人为行为不仅增加了输送到沿海系统的 N 总量,还增加了 N:P 的比率(Nixon 1993,Benitez-Nelson 2000)。这些对 N:P 比率的扰动在整个生态系统中并不统一,表明受影响更严重的生态系统,那些通常具有较高 N:P 比率的生态系统,可能会产生更多的 N2O。 还会抑制潮间带沉积物从大气中去除 N2O 的能力,可能导致 N2O 排放量增加。
生态系统 N2O 动态
为了估计我们测量的 N2Ofluxes 对这个系统的潜在影响,我们可以将它们放大到生态系统水平。作为近似值,我们可以使用我们在 6.7 6 2 lmol m-2 h 的实地研究中测量的平均通量—1,并且,假设这个平均值在空间上代表了梅岛河口生态系统中 7.2 平方公里的潮汐,我们计算了夏季(6 月、7 月和 8 月),暴露的潮汐为 106 kmol N2O。然后,使用一个范围在温带潮下带河口沉积物中测量的 N2Oflux 的数量(Seitzinger 和 Nixon 1985,Dong 等人 2002)并将这些值应用于 Plum Island Sound 河口区域,我们可以计算出可能的夏季 N2Ofluxes 范围约为 26 至 3000 kmol。附近纳拉甘西特湾的 N2O 释放量落在这些值的较低端,当应用于 Plum Island Sound 区域时,可能会从 Plum Island Sound 河口产生 24.5 kmol 的夏季 N2O 排放量。因此,在夏季,潮滩作用作为一个单独可能更多的水槽 平衡的河口 N2O 产量基于来自附近的大型河口的估计。我们还可以包括来自梅岛海峡盐沼边缘的 N2Oflux。使用 33 lmol m-2 d-1 的夏季 N2Oflux(Moseman-Valtierra 等人,2011 ) 应用于生态系统中 39 km2 的盐沼,我们得到了 117 kmol N2O 被沼泽吸收的夏季通量。计算夏季净生态系统吸收 200 kmol N2O。这是该系统提供的从大气中去除 N2O 的大量生态系统服务,被归类为一组被认为是相反的系统(Seitzinger 等人,2007)。 2000,Kroeze 等人,2005 年。此外,这是沿海系统提供的又一项重要服务,受人为活动危害。增加 N 输送,也许更重要的是,由于人类活动导致沿海系统 N:P 比率增加阻碍的潜力 r 潮汐滩从大气中去除这种强效温室气体的能力。
潮汐脉冲对于温和潮间带泥滩中一氧化二氮通量改变的研究——摘要、介绍
潮汐脉冲对于温和潮间带泥滩中一氧化二氮通量改变的研究——方法、数据分析
潮汐脉冲对于温和潮间带泥滩中一氧化二氮通量改变的研究——结果
