热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
结果和讨论
同时进行偶氮还原和电流生成。 在MFC或NA反应器中,苋菜红在28小时内完全还原(图1A),但含有10 mM苋菜红的MFC除外,其中0.7 mM苋菜红因电子供体耗尽而残留。 在非生物反应器中未检测到苋菜红还原。 MFC中的苋菜红还原率通常低于含有相同苋菜红浓度的NA反应器中的还原率,这表明电流的产生使一些电子流偏离了偶氮还原。 此外,所有反应器中的苋菜红还原率随着苋菜红浓度的增加而增加,尽管我们研究中使用的偶氮染料浓度(高达10 mM)高于其他报告中使用的浓度,31表明该菌株具有较高的偶氮还原能力。
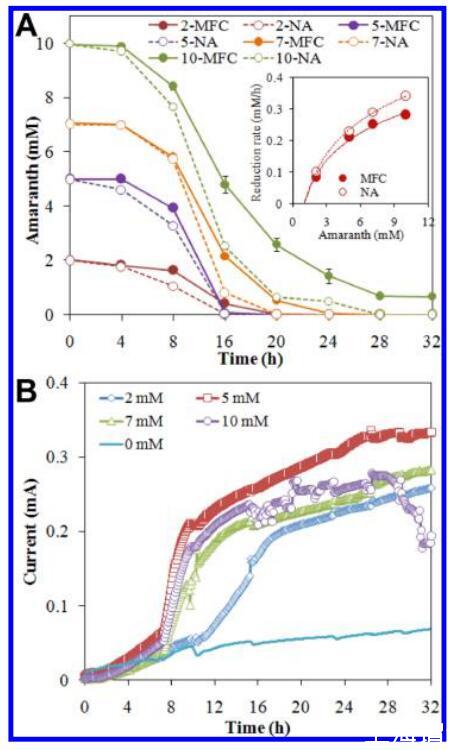
图1。 同时减少苋菜红和电流产生。 (A) 在MFC(实线)和NA反应器(虚线)中还原不同浓度的苋菜红。 插图显示了MFC和NA反应器中苋菜红的还原速率。 (B) 含有不同浓度苋菜红的MFC中的当前世代。
图1B显示了当前一代带有或不带苋菜红的MFC(完整的当前一代周期如图S3所示,支持信息)。 苋菜MFC在接种时产生0.01 mA的电流,而带有苋菜红的MFC在电流产生之前有大约8 h的滞后期(图1B),这表明浮游细胞更喜欢苋菜红作为电子受体而不是阳极。 伴随着偶氮还原,含有苋菜红的MFC的电流在滞后阶段后迅速增加,几乎是不含苋菜红的MFC的2倍(即,含有2、5、7和10 mM苋菜红的MFC的电流分别为0.3、0.34、0.29和0.28 mA,而不含苋菜红的MFC的电流为0.13 mA)。 在含有苋菜红的MFC中产生的增强电流可归因于苋菜红还原产生的萘酚的电子介体作用。含有5 mM苋菜红的9个MFC产生的电流最高, 由于毒性,在较高苋菜红浓度下观察到脱色S.细胞的代谢活性降低(图S4,支持信息)。32因此,在后续实验中,5 mM苋菜红被用作替代电子受体。
浮游生物和生物膜细胞生长。 含有苋菜红的MFC中基于蛋白质的浮游生物量约为无苋菜红MFC的两倍,但与具有相同苋菜红浓度的NA反应器中的浮游生物量无显着差异(图2A)。 结果表明,阳极呼吸对浮游细胞的生长没有显着影响,即浮游细胞的生长主要由苋菜红还原而不是电极呼吸驱动。 这与先前的报告一致,即脱色链球菌当前的产生是以生物膜为主,阳极呼吸的脱色链球菌细胞更倾向于附着在阳极表面,而不是悬浮在培养液中。 33,34 含有苋菜红的MFC中基于蛋白质的浮游生物量约为无苋菜红MFC的两倍,但与具有相同苋菜红浓度的NA反应器中的浮游生物量无显著差异(图2A)。结果表明,阳极呼吸对浮游细胞的生长没有显著影响,即浮游细胞的生长主要由苋菜红还原而不是电极呼吸驱动。这与先前的报告一致,即脱色链球菌当前的产生是以生物膜为主,阳极呼吸的脱色链球菌细胞更倾向于附着在阳极表面,而不是悬浮在培养液中。33,34
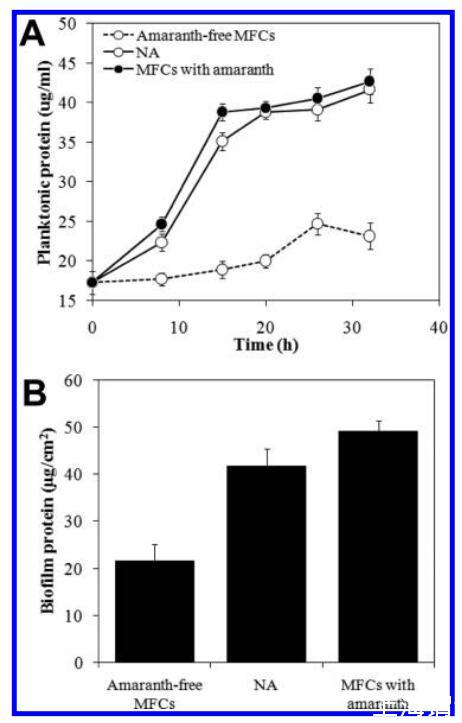
图2。 脱色链球菌的生长受不同电子受体的影响。 (A) 蛋白质浓度的变化,作为有无苋菜红(5 mM)或钠反应器的MFC中浮游细胞的指标。 (B) 32小时后,带有或不带有苋菜红或钠反应器的MFC阳极上的蛋白质含量。
与浮游细胞不同,含有苋菜红的MFC中阳极生物膜的蛋白质含量显着高于不含苋菜红的MFC(P<0.01)和NA反应器(P<0.05)(图2B),表明偶氮还原和电流生成都有助于生物膜的生长。 生物膜生物量的差异表明,在含有苋菜红的MFC中,偶氮还原占生物膜生物量的主要部分,而当前一代占较小的部分。 这些生物量差异与基于吉布斯自由能计算的预期差异一致,这表明苋菜红还原产生的自由能可能高出3倍以上( 生物膜生物量的差异表明,在含有苋菜红的MFC中,偶氮还原占生物膜生物量的主要部分,而当前一代占较小的部分。这些生物量差异与基于吉布斯自由能计算的预期差异一致,这表明苋菜红还原产生的自由能可能高出3倍以上(−61.8 J,用公式S1计算,支持信息)比阳极呼吸产生的值高(−17.7 J,用等式S2计算,支持信息),在含有5 mM苋菜红的MFC中。
生物膜中的pH和氧化还原电位曲线。 用O2微电极进行的测量证实,在实验过程中,散装液体是缺氧的(图S5,支持信息)。 pH微电极测量(图3A,B)表明,随着细胞生长,体积液体pH值降低,即,含有苋菜红的MFC(6.43)<NA反应器(6.55)<无苋菜红的MFC(6.65),表明谢瓦纳菌细胞外呼吸期间分泌酸性代谢物。 由于细菌电极呼吸是一种质子释放反应,因此含苋菜红的MFC中电流产生的更大增加也会降低pH值。 在无苋菜红的MFC的生物膜中观察到pH梯度下降(约0.02),这表明质子在阳极呼吸生物膜中的积累与之前的预测一致。35本研究中观察到的pH梯度小于在其他阳极呼吸生物膜中观察到的pH梯度(0.08到1个单位), 17,36这可能是由于较薄的生物膜或Shewanella生物膜相对较高的扩散效率。37与阳极呼吸生物膜相比,分别与阳极和苋菜呼吸或仅与苋菜呼吸时,生物膜中的pH梯度降低或消失。 这是因为质子可能被苋菜红还原生成的萘胺所消耗。
用氧化还原微电极测定了不同生物膜内的氧化还原电位。 无细胞反应器的结果表明,苋菜红对氧化还原电位曲线没有任何显着影响(图S6,支持信息)。 所有接种反应器的散装液体中的氧化还原电位从阳极表面上方约4mm处向阳极急剧下降(图3C),表明还原当量的累积在生物膜之外的可测量距离(几毫米)处开始。 无论使用何种电子受体,所有生物膜内的电位持续降低约30 mV(图3C,D)。 Shewanella细胞和电极之间的电子转移基本上是由黄素作为外膜细胞色素c(如MtrC/OmcA)的辅因子介导的,这已在菌株S12.12,24,38催化的MFC中得到证实。 生物膜中的pH变化可能导致1.2或2.4 mV的电位梯度 17苋菜红呼吸生物膜电极表面电位下降是预期的,因为电子从生物膜转移并逐渐氧化为散装液体,如生物膜呼吸中所观察到的 39,40 Picioreanu及其同事预测,含有电子介体的阳极呼吸生物膜内的氧化还原电位会随着电子介体的氧化还原状态先降低,然后升高。 35该预测适用于Shewanella生物膜,正如Shewanella阳极呼吸一样 38阳极呼吸生物膜内的电位持续下降至 17 无细胞反应器的结果表明,苋菜红对氧化还原电位曲线没有任何显著影响(图S6,支持信息)。所有接种反应器的散装液体中的氧化还原电位从阳极表面上方约4mm处向阳极急剧下降(图3C),表明还原当量的累积在生物膜之外的可测量距离(几毫米)处开始。无论使用何种电子受体,所有生物膜内的电位持续降低约30 mV(图3C,D)。Shewanella细胞和电极之间的电子转移基本上是由黄素作为外膜细胞色素c(如MtrC/OmcA)的辅因子介导的,这已在菌株S12.12,24,38催化的MFC中得到证实。生物膜中的pH变化可能导致1.2或2.4 mV的电位梯度(分别针对一个或两个电子转移过程,使用公式S3计算,支持信息),其远小于检测生物膜中的电位梯度(30 mV)这与Shewanella阳极生物膜电位下降与pH无关的建议一致。17苋菜红呼吸生物膜电极表面电位下降是预期的,因为电子从生物膜转移并逐渐氧化为散装液体,如生物膜呼吸中所观察到的与氧、硫酸盐或硝酸盐混合。39,40 Picioreanu及其同事预测,含有电子介体的阳极呼吸生物膜内的氧化还原电位会随着电子介体的氧化还原状态先降低,然后升高。35该预测适用于Shewanella生物膜,正如Shewanella阳极呼吸一样受到电子介质,即黄素的抑制。38阳极呼吸生物膜内的电位持续下降至−然后在阳极表面几微米(<10μm)范围内,340 mV显著增加至102 mV,这与上述预测一致,并且与使用极化阳极呼吸的S.oneidensis MR-1生物膜中的电位分布相似。17

图3.不同反应器中阳极上方的pH和氧化还原电位分布。(A)从液体表面到阳极表面的pH分布。(B)阳极表面上方95μm范围内pH分布的放大。(C)从液体表面到阳极表面的氧化还原电位分布。(D) 阳极表面上方95μm范围内的电位分布放大。0μm深度代表生物膜 − 阳极界面,虚线表示近似的生物膜 − 液体界面(红色表示苋菜呼吸生物膜;绿色表示阳极呼吸生物膜;蓝色表示生物膜同时呼吸),通过实时显微镜监测(300×)和CLSM生物膜结构观察确定界面。
阳极表面电位( −含有苋菜红的MFC和不含苋菜红的MFC(分别为68或102 mV)远高于相应的生物膜电位。此外,可通过改变阴极条件来调节阳极表面电位,即断开外部电路或降低阴极铁氰化物浓度可显著降低MFC阳极表面电位,当连接钠反应器的阳极和阴极时,可能会导致阳极表面电位急剧增加,并立即产生电流(0.2 mA,图S7,支持信息)。然而,这种阴极变化并未对所有反应器中的生物膜电位产生直接影响,表明生物膜内的氧化还原状态不受影响。通过生物膜和阳极表面之间的电位间隙进行电子转移产生的能量将由细菌细胞而不是电流产生来保存。28因此,适当降低阳极表面电位可能会增加阴极−阳极电压和增加MFC电流的产生。Shewanella阳极呼吸生物膜内的电位下降趋势与地杆菌生物膜相反。基于生物膜电子载体(PilA、细胞色素c或黄素)和扩散效率的差异,在两种模型外生电子的生物膜内可以假设13种不同的电子转移机制。
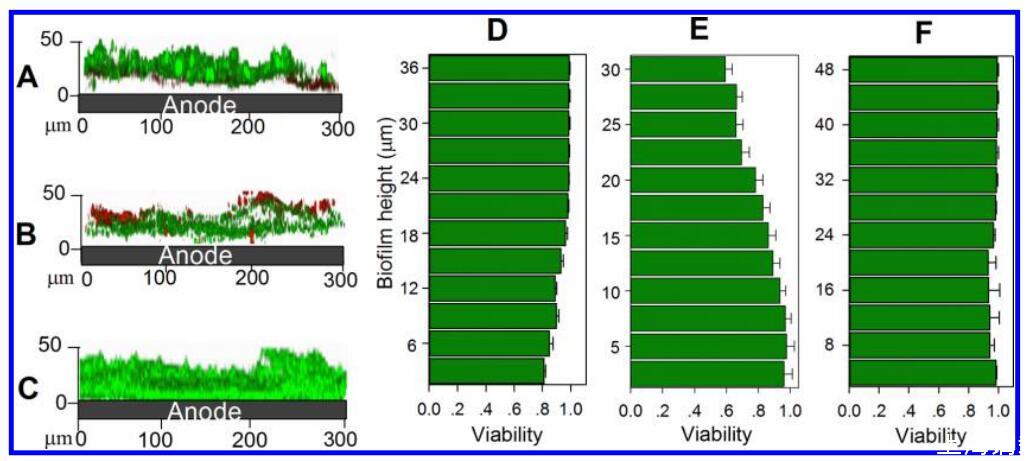
图4。 不同电子受体呼吸的生物膜内的活性分布。 A、 B和C分别是在CLSM下观察到的苋菜红呼吸生物膜、阳极呼吸生物膜和同时苋菜红和阳极呼吸生物膜的垂直截面。 D、 E和F分别是基于苋菜红呼吸生物膜、阳极呼吸生物膜和同时苋菜红和阳极呼吸生物膜的CLSM结果的定量活性曲线。 低活性和受损膜的细胞呈红色,而高活性和完整膜的细胞呈绿色。 0μm处的生物膜层为生物膜 − 电极界面(底部)。
生物膜内的活性分层。 在所有生物膜中均观察到不均匀分布的塔状细胞簇结构。 与生物膜蛋白含量的观察类似,含有苋菜的MFC中的生物膜(48μm)比苋菜MFC(32μm)或NA反应器(41μm)中的生物膜厚(图4)。 活性染色显示,阳极和苋菜红共同呼吸的生物膜总活性(0.97)高于仅阳极和苋菜红呼吸的生物膜(0.94)或苋菜红(0.66)。 仅用苋菜红呼吸的生物膜的活性从电极上增加 − 生物膜与生物膜的界面(底部,0.66) − 液体界面(顶部,0.96)。 该曲线与呼吸氧气或其他可溶性电子受体的生物膜中的曲线一致。22,41相反,阳极呼吸生物膜中的活性分层从底部(0.99)下降到顶层(0.81),类似于S.oneidensis MR-1、G.sulfurreducens、, 铜绿假单胞菌和革兰氏阳性细菌Thermincola potens。 23,42,43可以用生物膜中的电子跳跃电子转移机制来解释,无法接近阳极表面的细菌细胞不能以足够的速率将电子转移到阳极, 这将生物膜细胞的细胞代谢活性限制在阈值距离内。13,44这也可以解释为什么Shewanella阳极呼吸生物膜的厚度通常限制在几十微米以内。14相反, Geobacter阳极呼吸生物膜的厚度可以达到数百微米,并保持均匀的导电性,14尽管其扩散效率低于Shewanella生物膜,37进一步表明Shewanella使用不同的电子转移机制。 阳极呼吸生物膜底部的高活性也排除了苋菜呼吸生物膜中活性降低是由于质子积累或缺乏乳酸引起的可能性。 因此,仅使用阳极或苋菜红呼吸的生物膜内的反向活性曲线依赖于电子受体,并且与假定的电子转移曲线一致。 这在同时呼吸阳极和苋菜红的生物膜中有所不同,这表明在生物膜中具有更高的均匀活性, 表明生物膜内的双向电子转移可以补偿阳极或苋菜红作为唯一电子受体无法接近的生物膜细胞的活力损失。
生物膜的呼吸系统和生理层次中的电子受体的依赖性——摘要、介绍
生物膜的呼吸系统和生理层次中的电子受体的依赖性——材料和方法
 相关新闻
相关新闻