热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
温度和沉积层对沉积物中不同种类微囊藻菌落存活的影响
微电极分析表明,含有微囊藻菌落的表层为需氧层,深层为厌氧层(支持材料中的图S1)。
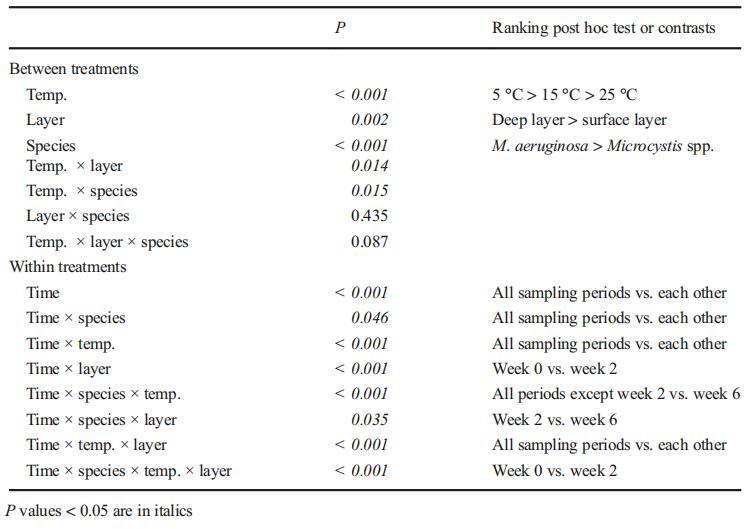
微囊藻菌落在不同沉积条件下的存活能力如图2所示。通过三因素重复方差分析,分析了温度、微囊藻种类和沉积层对微囊藻菌落存活的影响。温度是显着影响底栖微囊藻菌落存活的唯一因素(P<0.001,表1)。具体而言,25°C的温度对长期存活不利,如相应的事后测试所示。此外,存在显着的温度-层相互作用(P=0.003),表明层间差异取决于所讨论的温度,反之亦然。三次重复的单因素方差分析显示,在15°C(P=0.007)和25°C(P<0.001)时,沉积物深层微囊藻菌落的存活率显着大于表层微囊藻菌落的存活率,而在5°C时,微囊藻菌落的存活率无显着差异(P=0.260)。在5°C时,卫森伯格支原体的存活率显着高于铜绿支原体(P=0.018)。然而,在15°C和25°C时,卫森伯格支原体和铜绿支原体的存活率没有统计学差异(分别为P=0.740和0.981)。
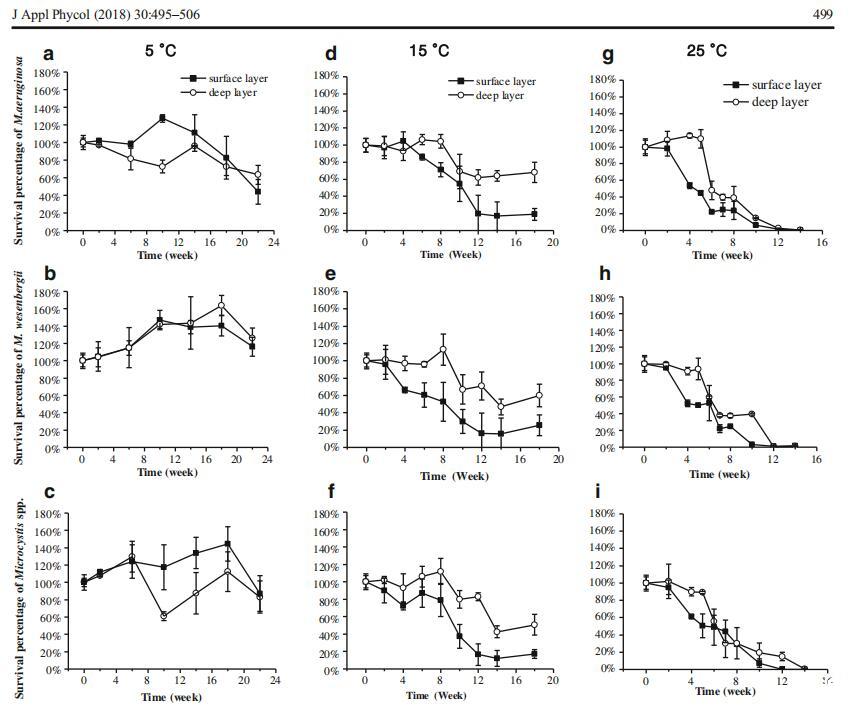
图2不同温度下沉积物表层和深层微囊藻菌落的存活率,表示为存活百分比:5℃下铜绿微囊藻a、5℃下魏森伯格微囊藻b、5℃下微囊藻C、15℃下铜绿微囊藻d、15℃下魏森伯格微囊藻e、15℃下微囊藻f、25℃下铜绿微囊藻g,h M.wesenbergii在25°C下,i.微囊藻在25°C下。根据我们的实验前结果,针对不同的温度设置了不同的采样间隔
如表1所示,时间的处理内效应以及时间与三个因素(温度、微囊藻种类和层数)之间的关系都是显着的,表明沉积物中存活微囊藻细胞的密度随时间而变化。此外,变化模式因微囊藻种类、温度和沉积层而异。因此,通过单因素重复方差分析和Tukey事后检验,分别研究了不同沉积条件下不同微囊藻群的变化规律。
表1重复测量了14周内温度(5、15和25°C)和沉积层(表层和深层)对不同种类微囊藻菌落(铜绿假单胞菌、魏森伯格微囊藻和这两种微囊藻的1:1混合物)存活率影响的方差分析
在5°C温度下,沉积物表层和深层中存活的铜绿假单胞菌的丰度在前18周内没有显着变化,仅在第22周下降(分别为P=0.048和0.044)。在整个实验过程中,表层魏森伯格支原体和微囊藻的丰度没有显着差异(分别P=0.134和0.191)。在深层,在整个实验过程中,细胞数量具有可比性,但卫森伯格支原体在第18周(P=0.020)和混合微囊藻菌落在第10周(P=0.003)除外。当比较初始(第0周)和最终(第22周)细胞密度时,仅发现表层中的铜绿假单胞菌存在显着差异。
在15°C下,在初始滞后期后,表层微囊藻丰度的减少变得相当迅速,此后从第12周到试验期结束,存活率稳定在约25%。铜绿微囊藻的滞后期为4周(第4周与第6周的P值为0.015),而威森伯格微囊藻的滞后期为2周(第4周与第6周的P值为0.013)。在深层,所有三个系列的微囊藻菌落在前8周保持稳定,仅在第8周至第12周(铜绿假单胞菌)或第14周(维森贝格氏微囊藻菌落和混合微囊藻菌落)出现明显损失。随后,未发现显着下降,试验结束时,所有三个系列的微囊藻的存活率均保持在50%以上(铜绿假单胞菌为64.58%,韦森贝基微囊藻为59.22%,混合微囊藻菌落为50.80%)。
在25°C下,所有三个微囊藻系列的动力学表现出相似的趋势。经过2周的滞后期后,表层微囊藻的存活率降至接近零,直到第12周。在深层,5周的滞后期显着延长。铜绿假单胞菌和混合微囊藻菌落的存活率在第14周降至接近零,而卫森伯格假单胞菌的存活率在第12周降至接近零。
底栖微囊藻群体中微囊藻毒素总含量和微囊藻毒素配额的动态
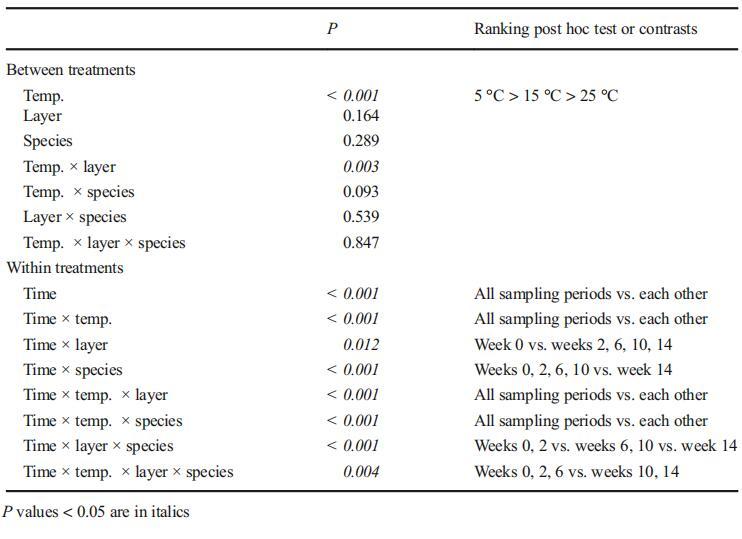
计算并比较了沉积物中微囊藻菌落的总MC含量。MCLR和MC-RR的浓度在所有条件下均表现出良好的相关性(皮尔逊相关性,N=192,R=0.980,P<0.001)(图3)。因此,在本研究中,我们没有将它们分开,而是将它们统称为BMC^。我们比较了三组微囊藻菌落的存活率:毒性铜绿假单胞菌、无毒魏森伯格微囊藻菌落和两种微囊藻的1:1混合物(表示为微囊藻属)。仅分析了铜绿假单胞菌和微囊藻组的MC含量。MC含量的动态表现在两个层面:表层或深层沉积物中所有存活微囊藻菌落的MC总含量和MC配额,其计算方法为总MC含量除以细胞计数的商。微囊藻群的MC配额可能受到铜绿假单胞菌和威森伯格假单胞菌丰度比的影响。因此,仅使用铜绿假单胞菌的MC配额来反映每个细胞间MC含量的变化(图4)。
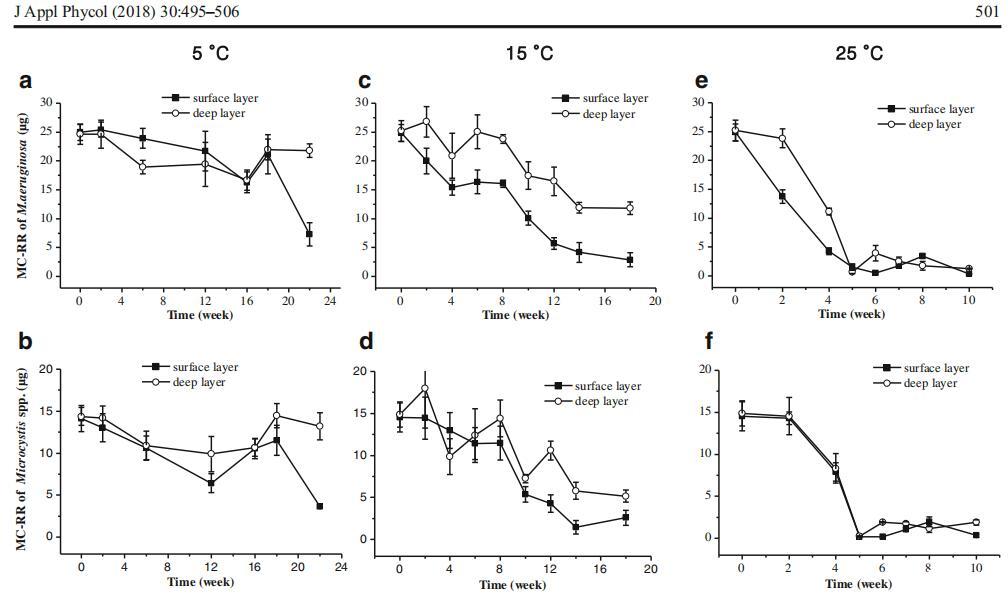
图3 5°C下铜绿假单胞菌a和微囊藻PP沉积物表层和深层微囊藻菌落中MC-RR总含量的动态。在5°C下,铜绿假单胞菌在15°C下,d微囊藻PP。在15°C下,铜绿假单胞菌在25°C下,微囊藻属在25°C下。根据我们的实验前结果,针对不同的温度设置了不同的采样间隔
图4 5°C(a)、15°C(b)和25°C(C)条件下沉积物表层和深层中铜绿假单胞菌的微囊藻毒素配额。根据我们的实验前结果,针对不同的温度设置了不同的采样间隔
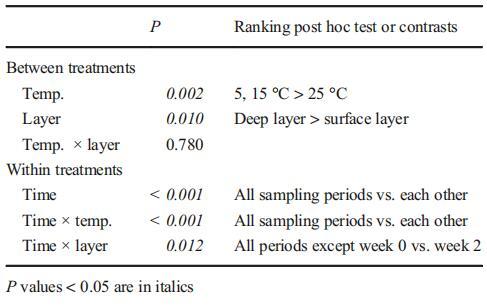
温度和沉积层显着影响沉积物中微囊藻菌落的总MC含量(表2)。在较低温度下,微囊藻菌落的总MC含量在深层显着较高。此外,存在显着的温度-层相互作用(P=0.003),表明层间差异因温度而异。
表2重复测量温度(5、15和25°C)和沉积层(表层和深层)对微囊藻菌落(铜绿假单胞菌、1:1的铜绿假单胞菌和韦森伯格假单胞菌混合物)MC-RR总含量影响的方差分析
在不同温度下分别进行单向重复方差分析和Tukey的事后检验。在5°C下,整个试验期间,铜绿微囊藻和混合微囊藻菌落中的MC总含量没有显着差异(表层铜绿微囊藻,P=0.062;表层微囊藻属,P=0.065;深层铜绿微囊藻,P=0.137;深层微囊藻属,P=0.087)。通过两两比较,第22周表层菌落的总MC含量显着低于其他采样日期。在15°C和25°C时,铜绿假单胞菌中的总MC含量在实验期间显着下降(P<0.05)。在15°C的深层,铜绿假单胞菌和微囊藻菌落的总MC含量在前8周保持稳定(铜绿假单胞菌,P=0.167;微囊藻,P=0.088),并从第10周到实验结束显着下降。在15°C的表层中,MC的总含量在第二周或第四周开始显着降低,在其余的实验期间,这种降低趋势持续。在25°C时,铜绿假单胞菌和微囊藻菌落的总MC浓度在前5周迅速下降,随后在表层和深层均保持较低水平。表层铜绿假单胞菌细胞中MC浓度显着低于深层铜绿假单胞菌(P<0.001)。
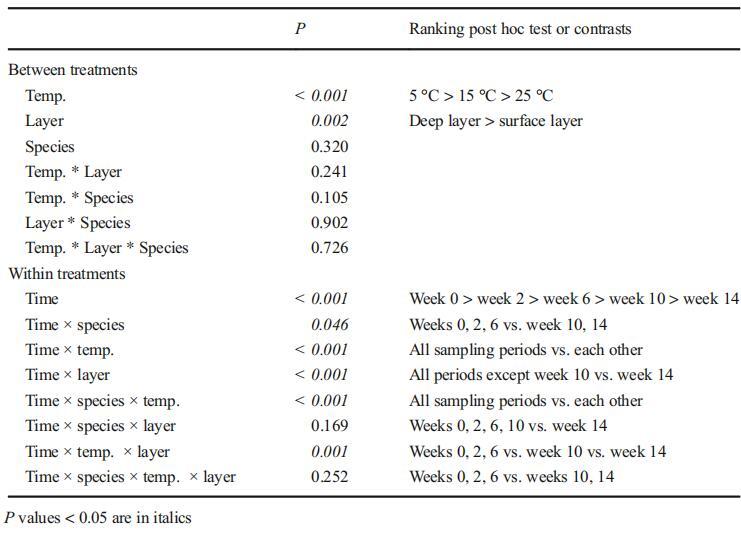
温度和沉积层显着影响绿脓杆菌的MC配额(P<0.001,表3)。在较高温度(25°C)和表层,铜绿假单胞菌的微囊藻毒素配额显着降低。时间的内处理效应,以及时间与温度和层之间的关系都是显着的。
表3重复测量了14周内温度(5、15和25°C)和沉积层(表层和深层)对铜绿假单胞菌微囊藻毒素配额影响的方差分析
对每个温度进行单因素方差分析和Tukey的事后检验。与三因素方差分析的统计结果不同,根据单因素方差分析(5°C,P=0.062;15°C,P=0.449;和25°C,P=0.371),两层沉积物中铜绿假单胞菌的MC配额之间未发现显着差异。在5°C时,第一周铜绿假单胞菌的MC配额与第6、第10和第22周的MC配额相当,但显着低于或高于第2、第14和第18周的MC配额。在15°C时,只有第2周和第14周的MC配额与初始值显着不同。在5°C和15°C下采集的样品中,铜绿假单胞菌的MC配额没有显着差异。虽然在一些采样日期存在波动,但微囊藻毒素配额在5°C和15°C下在表层和深层沉积物中基本稳定。在25°C下,任何采样日期的MC配额都明显低于实验开始时。
沉积物中微囊藻群体光合活性的动态
使用三因素重复方差分析测试分析温度、微囊藻种类和层数对微囊藻菌落Fv/Fm的影响(图5)。温度和沉积层显着影响底栖微囊藻菌落的Fv/Fm(P<0.01,表4)。在所有沉积条件下,微囊藻的Fv/Fm逐渐降低,在较高温度(P<0.001)和表层(P=0.002)下,Fv/Fm的下降百分比增加。毒性铜绿假单胞菌和无毒韦森伯格假单胞菌的Fv/Fm无显着差异(P=0.320)。
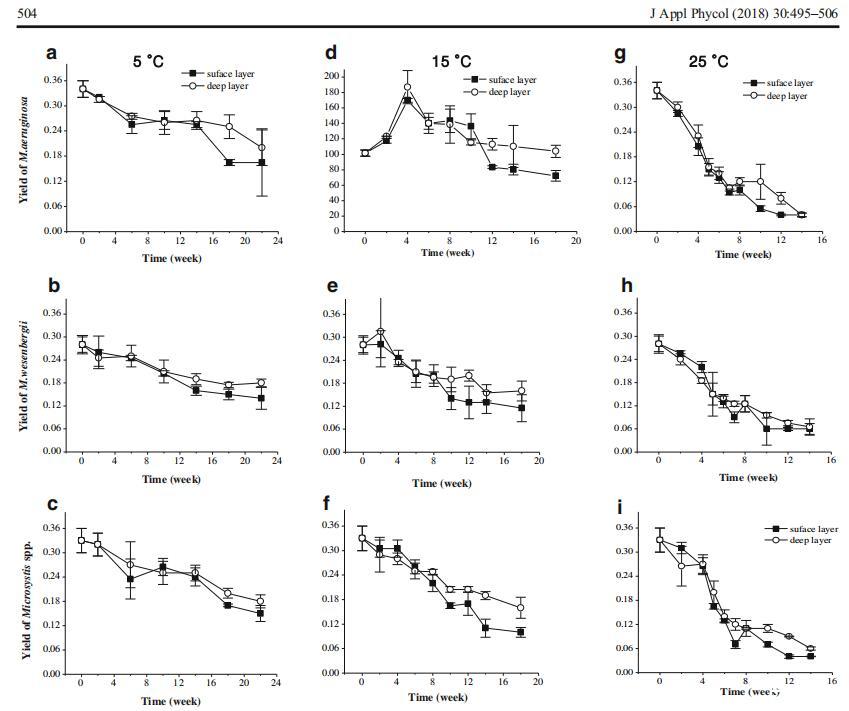
图5不同温度下沉积物表层和深层微囊藻的有效量子产率(单位:PSII(Fv/Fm))。铜绿假单胞菌在5°C时,铜绿假单胞菌在5°C时,铜绿假单胞菌在5°C时,铜绿假单胞菌在15°C时,铜绿假单胞菌在15°C时,铜绿假单胞菌在15°C时,铜绿假单胞菌在25°C时,铜绿假单胞菌在25°C时,铜绿假单胞菌在15°C时,铜绿假单胞菌在15°C时,铜绿假单胞菌在15°C时,铜绿假单胞菌在25°C时,铜绿假单胞菌在25°C时,铜绿假单胞菌在25°C时,铜,根据我们的预实验结果
表4重复测量了14周内温度(5、15和25°C)和沉积层(表层和深层)对绿脓杆菌光合活性影响的方差分析
治疗时间内的疗效显着。微囊藻菌落的丰度开始下降之前有一段滞后期。然而,在整个实验过程中,Fv/Fm值显着降低。时间与三个因素(温度、微囊藻种类和层)之间的关系显着,表明下降速度受微囊藻种类、温度和沉积层的影响。
在这项研究中,我们考察了在受控实验室条件下,考虑温度和氧气的影响,底栖微囊藻菌落的存活能力。我们不打算在不同温度下模拟底栖微囊藻群落的整个寿命,因为浅水湖泊中沉积物的温度是周期性的,并随季节变化。相反,我们采用了为期4-6个月的模拟试验来评估底泥中微囊藻菌落的存活能力。
在5°C条件下,表层和深层沉积物中的微囊藻菌落丰度在实验结束时(第22周)没有显着减少,表层中的铜绿假单胞菌除外。在15°C和25°C时,沉积物中微囊藻菌落的丰度在初始滞后期后迅速下降,随后存活率稳定下来,直到实验结束。物流模型(方程式1)大致符合沉积物中微囊藻菌落在15°C和25°C下存活百分比的动态:
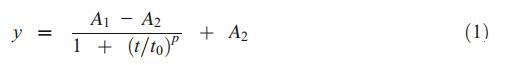
其中t、y、t0和p分别代表采样时间、沉积物中微囊藻的存活百分比、中心时间和功率。A1和A2代表存活率稳定的初始值和最终值。通过对模型进行一阶导数,得到了变化过程的开始时间和结束时间,分别用t1和t2表示。也就是说,微囊藻细胞的存活率在t1的初始滞后期后开始下降,此后存活率从t2到试验期结束稳定在A2(表5)。
表5根据逻辑模型计算的15或25°C条件下不同沉积物层中微囊藻菌落的存活百分比
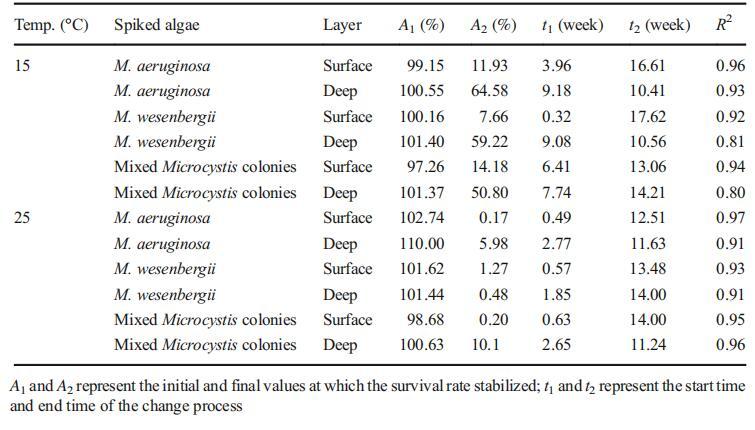
较大的初始延迟时间(t1)和最终存活率(A2)可以反映较高的存活率。由于在5°C下22周的实验过程中几乎没有发现任何显着的下降,t1和A2可分别被视为22周和100%。通过比较不同条件下微囊藻菌落的t1和A2,我们得出以下结论:(1)低温有利于微囊藻菌落在沉积物中更长的存活时间,(2)铜绿假单胞菌的最终存活百分比(表5中的A2)在15℃和25℃时高于卫森伯格微囊藻,深层微囊藻的存活率显着高于表层微囊藻。微囊藻菌落的存活能力可与植物种子的存活能力相比较,植物种子埋在土壤中时,可作为下一季萌发的种子库;硬壳种子可以在土壤中存活数十年,同时在适当条件下保持发芽能力。因此,沉积微囊藻菌落也可以被认为是一种种子库;在野外条件下,微囊藻形成由数十个、数百个或数千个细胞组成的菌落,细胞被厚厚的鞘覆盖。就像种子的外壳一样,微囊藻菌落的鞘可以保护微囊藻细胞免受不利的底栖条件的影响。低温和厌氧条件促进了种子和底栖微囊藻群体的长期存活(Andrade,2001年)。
温度是影响沉积物中微囊藻菌落存活的最重要因素。在5°C温度下22周的实验过程中,微囊藻数量几乎没有显着减少,而在25°C温度下,超过90%的底栖生物种群在3个月内消失。这些结果与现场调查结果一致。例如,微囊藻菌落可以在格兰根特湖(法国)的沉积物中存活长达6年,那里的温度总是低于10℃(Misson等人,2012a)。在德国Quitzdorf水库的另一个湖泊的沉积物中,当温度在再侵入阶段持续高于12°C时,65%到85%的底栖微囊藻群落死亡(Ihle等人,2005年)。本研究定量揭示了沉积物中微囊藻菌落的存活情况。根据我们的研究结果和亚热带浅水湖泊温度的实际季节变化,大多数底栖微囊藻群落能够成功越冬,因此有可能在第二年春天接种水柱,但它们不能在亚热带浅水湖泊的沉积物中积累多年。
氧气水平是决定底栖微囊藻菌落存活的另一个因素。微囊藻菌落在好氧表层沉积物中的存活率显着低于厌氧深层沉积物。如表1所示,在15°C下放置17周后,上层沉积物中微囊藻菌落的丰度下降,存活率低于20%,而深层沉积物中的种群减少率低于50%。其他研究人员也报告说,沉积物表层中有更高比例的微囊藻菌落消失(Brunberg和Blomqvist,2002年;Ihle等人,2005年),这可能是再侵入和细胞溶解的结果。其他蓝藻,如圆柱状振荡藻,也对氧敏感,在还原条件下比有氧条件下存活更长(Richardson和Castenholz 1987)。
毒性铜绿假单胞菌和无毒维森伯格假单胞菌具有相似的存活能力,尽管铜绿假单胞菌在15°C和25°C下的最终存活效率(表5中的A2)高于维森伯格假单胞菌。一些研究人员发现,MCs可能参与了底栖微囊藻的存活机制。因此,有毒微囊藻菌落可能比无毒菌落更容易越冬(Ihle等人,2005年;Misson等人,2012a)。然而,在我们的研究中,毒性铜绿假单胞菌和无毒维森伯格假单胞菌的存活率没有显着差异。覆盖鞘的厚度可能是影响微囊藻生存能力的关键因素。在沉积物中,有鞘光养生物比无鞘光养生物寿命更长(Jørgensen等人,1988年)。铜绿假单胞菌的鞘比维森伯格假单胞菌的鞘薄得多。因此,有毒铜绿假单胞菌中的MC可能有利于其长期存活。然而,这一优势可能被铜绿假单胞菌较薄的鞘层所抵消。在另一项研究中,在沉淀物中埋藏6年和33年的底栖微囊藻种群之间,未发现有毒和无毒细胞比例的显着差异(Rinta Kanto等人,2009年)。需要进一步的研究来揭示细胞内MC在底栖微囊藻菌落存活中的生理作用。
在5℃和15℃下,微囊藻细胞中的MC含量在实验期间基本保持稳定。因此,可以推断,在黑暗条件下,沉积物中不会产生MCs。此外,底栖微囊藻保存的MCs没有作为碳源消耗或释放到周围环境中。然而,在25°C时,微囊藻细胞的MC配额显着降低。在此温度下,微囊藻细胞的光合活性显着降低。微囊藻细胞膜也可能受损,细胞间MC可能从细胞中释放,导致MC配额下降。Wu等人(2015年)还发现,新鲜蓝藻样本中的细胞内MC在细胞破裂之前不会迅速减少。对于野生样本,微囊藻种群包括有毒和无毒两种形式。因此,现场样本中MC配额的波动可能主要取决于有毒和无毒细胞的相对比例,而不是有毒细胞的细胞内MC含量水平。在我们的研究中,定量测定了群体微囊藻菌株纯培养物中细胞内MC的动态,从而消除了群落组成的任何潜在影响,并提供了更具针对性的结果。底栖微囊藻群体的光合活性逐渐下降,在较高温度下下降幅度较大。我们的研究结果和其他研究结果表明,在没有光照的情况下,沉积物中PSII的光化学效率逐渐降低(Schöne等人,2010)。当温度低于15°C时,毒性细胞和无毒细胞的存活率没有显着差异,细胞内含量稳定。因此,在越冬和再入侵期间,底栖微囊藻种群的毒性没有显着变化。
在这项研究中,我们系统地研究了微囊藻菌落在表层和深层沉积物中,在不同季节对应的不同温度下的存活能力。低温和厌氧条件促进了底栖微囊藻菌落的长期存活。毒性铜绿假单胞菌和无毒魏森伯格假单胞菌表现出相似的存活能力,在实验期间,底栖微囊藻细胞的MC含量在5℃和15℃下保持稳定。因此,在越冬和再入侵期间,底栖微囊藻种群的毒性没有显着变化。我们的研究表明,大多数底栖微囊藻群体能够成功越冬,微囊藻毒素得以保存,光合活性得以维持,显着增加了水华形成的风险。因此,需要采取疏浚或覆盖沉积物等人工措施,以避免沉积物中累积微囊藻菌落的不利影响。在夏季,当亚热带浅水湖泊的温度较高(例如25°C)时,底栖生物种群的存活率在3个月内降至接近零。因此,底栖微囊藻种群不能在亚热带浅水湖泊的沉积物中积累多年。
中国国家自然科学基金(31270509和31370504)、中国水污染防治与治理重大科学技术计划(2013ZX07102005)、国家淡水生态与生物技术国家重点实验室(2016FB10)等国家科学基金资助项目(41561144008和)。作者感谢有害藻类团队的所有成员,特别是中国科学院水生生物研究所的李仁辉教授,感谢他们对微囊藻菌株的纯培养和仪器的帮助。我们还要对袁晓博士(中国科学院水生生物研究所分析测试中心)的仪器支持表示最深切的感谢。
 相关新闻
相关新闻