热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
方法评价
实验1-测试方法。氧内流和传感器稳定性的控制实验。低O2浓度下的耗氧量测定要求极高的精度,且装置完全不受外部和内部O2污染[31]。为了确定低O2水平下的耗氧量,我们开发了一个全玻璃培养瓶,易于接近传感器,其中唯一可能的O2进入是通过25 cm长的压力补偿毛细管和传感器与8 mm玻璃管之间的薄水膜(图1)。压力补偿管的宽度为2.5 mm,根据菲克第一定律(方程式S1,文本S1)计算的氧气流量导致平衡时的输入为0.05 nmol L-1 h-1,这是完全可以忽略的。从孵育开始,试管中的水与散装水中的O2浓度相同,如果扩散是唯一的运输方式,则在许多月内不会达到平衡[32]。然而,水的粘度确实会随着温度的升高而降低,因此可能需要在较高温度下使用内径较小的压力补偿管,以尽量减少湍流混合。除了湍流混合外,通过压力补偿管的输送也可能因温度变化而增加,从而导致管内的水移动。管道中的水流沿管壁流动较慢,因此任何水流都会促进混合。另一个入口点是STOX传感器和周围玻璃管之间的水膜。假设水膜厚度为0.05 mm,在0.3 cm长的管中,在平衡状态下,只有0.02 nmol L-1 O2 h-1通过扩散通过该膜。然而,与压力补偿管一样,我们不能排除湍流混合的可能性,例如由振动泵引起的湍流混合。因此,由于我们无法排除与大气开放连接中的各种混合,我们决定通过使用生物灭活的0.05 M HCl脱盐水进行对照实验,测试是否可能泄漏到培养瓶中,该脱盐水已煮沸、冷却,然后用N2气体脱气至低O2水平。瓶内的氧气浓度(n=4)在0.60小时内没有增加(图2),这证实没有氧气泄漏。另一方面,在孵化过程中,传感器信号略有下降。根据特定的传感器,我们记录到信号的减少(培养64小时后),范围为初始值的2.2%到21.3%(前5小时内信号的平均值)。因此,对于培养持续14小时的实验,应考虑约1.8%的信号平均漂移。漂移在很大程度上取决于传感器的特定特性,因此,仅使用在使用前经过彻底测试并保持极化数小时的传感器,可以将漂移降至最低。然而,传感器漂移似乎只导致测得的O2浓度下降的一部分。更详细地分析图2中的数据,似乎瓶子中氧气的减少:STOX 1、2、3和4分别为0.14、0.07、0.04和0.15 nmol L-1 h-1(图2),与初始氧气浓度没有强相关性。因此,似乎大部分减少可能是由于我们无法完全停止氧气消耗,无论是生物还是化学。在这里,我们在浓度和绝对速率范围内工作,控制实验很困难,但基于大气浓度下的低传感器信号漂移速率[19],如STOX 1(图2)中所观察到的,3天内信号漂移约三分之一的可能性也很小。因此,我们对孵化期间真实传感器漂移的最佳估计为每14小时2%。
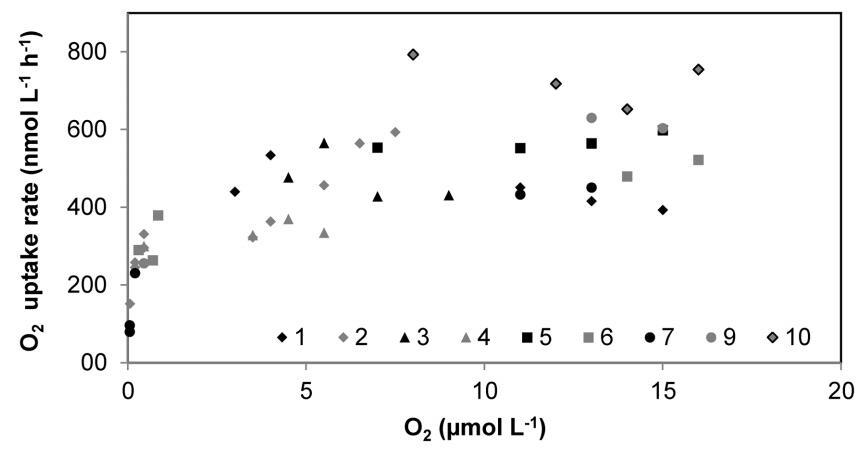
图4。氧气消耗率与1号站的氧气浓度(Randers峡湾,2011年9月)。每种类型的符号对应一个特定的瓶子。复制瓶在初始O2浓度范围内(从0到20 mmol L-1)孵育。速率计算为不同O2浓度变化间隔的斜率:每2 mmol L-1间隔为17-6 mmol O2,每1 mmol L-1间隔为6-1 mmol L-1 O2。然后在以下时间间隔内:500-300、300-100和100-5 nmol-1。内政部:10.1371/期刊。波内。0105399.g004
实验2-方法在沿海水域的应用和比较。通过测量丹麦沿海水域3个不同站点和3个不同季节的CR率,对该方法进行了实验测试,以覆盖群落组成和代谢水平的最小变化。本研究的两个主要站点(圣约翰海峡1号站和圣约翰海峡2号站,Marselisborg Marina,图S1)分别于6月、7月、9月和1月取样,并进行了三组反应堆试验。由于我们的方法允许随着时间的推移连续记录氧气浓度,因此我们能够以高分辨率而非少量点监测和分析氧气消耗过程(图3)。因此,可以评估反应器中氧气的减少是线性的,从高氧气浓度下降到氧气浓度#1 mmol L-1。在大范围O2浓度的培养过程中,呼吸速率似乎与矩形双曲线的一般形状相匹配(图4),并遵循Michaelis-Menten动力学。使用公式2(图5)通过曲线拟合确定参数Vmax和Km。表1报告了三个研究期间St.1和St.2的估算Vmax值和表观半饱和浓度Km。
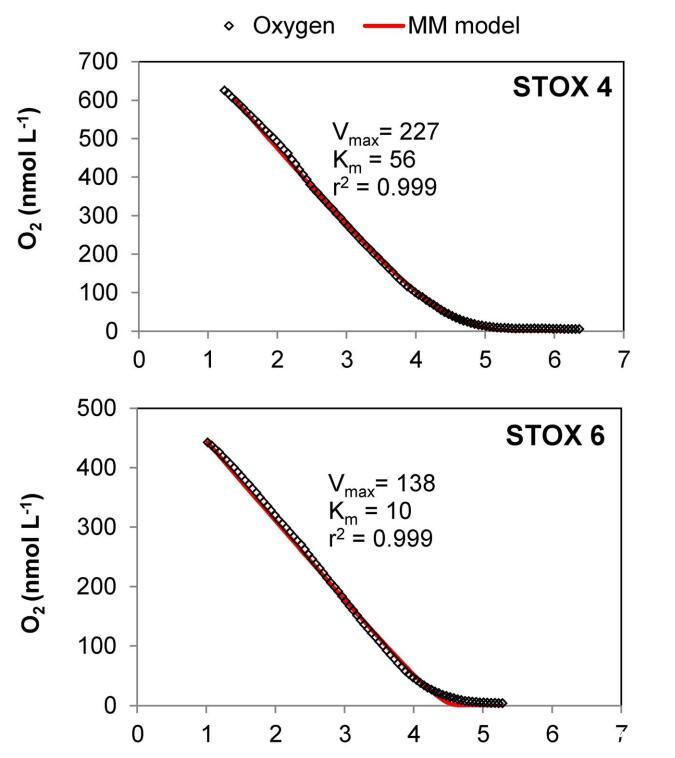

图5。沿海海水样品中氧气消耗曲线的建模示例。2011年9月,来自圣二号码头(Marselisborg Marina)的三个培养瓶(4、6、7)的耗氧实验。氧气耗尽后(6小时),将空气饱和水注入“STOX 7”瓶中。点显示STOX传感器记录的O2值。最大速率(Vmax)和半饱和常数(Km)通过最小二乘拟合(红色实线)确定,使用公式(2),源自Michaelis-Menten动力学。该方法的拟合度相对较差,可能导致低估“STOX 6”的Km值。内政部:10.1371/期刊。波内。0105399.g005
在氧浓度降低时,通过STOX传感器法测量的呼吸率和在空气饱和时通过温克勒滴定法测量的呼吸率得出了类似的结果。事实上,这两种方法记录的氧气吸收率大致相同,只要装有STOX传感器的瓶子中的氧气浓度高于500-200 nmol L-1,这取决于不同的群体(图6)。低于这些值时,呼吸速率取决于O2浓度,表明群落正在接近其表观Km值。2013年冬季,对北海的St.1和St.2以及额外的St.3进行了方法比较,结果相似(图6)。来自St.1、St.2和St.3的样品暴露于2–0.5 mmol L-1的O2浓度下,其吸氧率与暴露于285 mmol L-1 O2的样品相似(在标准误差范围内)。
因此,分别使用STOX传感器和Winkler滴定法进行测量,得出以下呼吸速率6标准误差(n):对于St.1,196.869(3)nmol L-1 h-1和203.8627(3)nmol L-1 h-1;对于St.2,163.562(13)nmol L-1 h-1和172.768.5(4)nmol L-1 h-1;对于St.3,98.664.9(6)nmol L-1 h-1和117.7613.5(4)nmol L-1 h-1。对于St.2浮游生物群落,在较低O2浓度(#500 nmol L-1)下进行的孵化似乎在略低于暴露于较高O2浓度(#2 mmol L-1)下的孵化达到最大速率(图6 D和F)。
耗氧和随后注入空气饱和水后测定的表观Km值与6月和9月St.1第一次耗氧曲线所得值相似。具体而言,对于6月的St.1,第一次消耗曲线的平均视在Km值6 St.误差(n)为83611(3)nmol L-1 O2,在添加约1 mmol L-1 O2及其后续消耗后,视在Km值为118625(3)nmol L-1 O2。9月,注入约0.5 mmol L-1 O2后,表观Km值首先为73625(3),然后为85626(3)nmol L-1 O2。然而,对于9月的St.2,第二次耗氧曲线导致表观Km值从第一次运行的179增加到第二次运行的310 nmol L-1 O2(表1和图5“STOX 7”)。
表1。测量的社区呼吸率概述。

只有当瓶子中的氧气浓度达到纳摩尔范围时,St.1和St.2的呼吸速率才会显着降低,对应于259-90 nmol-1 O2之间的氧气呼吸微生物群落的表观Km值。在夏季/秋季/冬季,这两个地点的估算Vmax值不同(表1),在186和761 nmol-1 h-1之间变化。通过根据从Jassby和Platt经验方法得出的数学函数(等式6)对数据进行建模,获得了Km和Vmax的类似值,但总体而言,动力学参数稍低,从而更好地描述了实验数据(图S3)。使用这种方法,估计的Vmax值比使用Michaelis-Menten模型获得的值更符合实际测量的呼吸率(CR)。这两个位置显示出相同的CR季节模式:夏季/秋季/冬季:605/262/223 nmol h-1(St.1)和238/206/156 nmol L-1 h-1(St.2),Km值在155-66 nmol L-1 O2之间变化。叶绿素a浓度(叶绿素a–表1)显示,浮游植物群落的丰度随季节的变化而变化,在St.1和St.2之间也有变化。截至目前,2011年6月Randers峡湾(St.1)样本中的chl a浓度最高,其呼吸速率也远高于其他样本(表1)。发现St.1的chl A浓度和CR之间存在线性相关(r2=0.99)。
实验3-海洋水域中的方法应用。该方法通过测量船上实验中的铬率,在海洋水样上进行了测试。水样在不同的瓶中以不同的氧气水平孵化(图7)。在培养14小时后,在三个重复瓶中检测到线性氧气下降。重复试验中的呼吸率相似,平均值为7.460.3(3)nmol L-1 h-1,与暴露时的O2水平(950/270/100 nmol L-1)无关。这表明该群落的Km值明显低于100 nmol L-1。

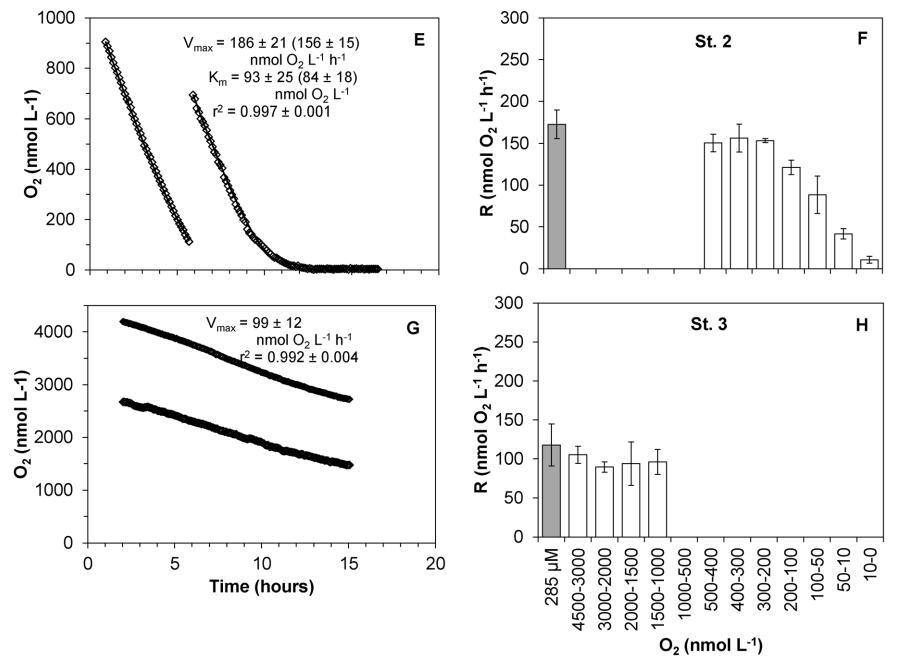
图6。用STOX法和温克勒滴定法测量铬率的比较。通过STOX法(从0到2 mmol L-1,白色条)和温克勒滴定法(在空气饱和时,灰色条)测量不同O2浓度(B、D、F、H)下培养期间的耗氧时间过程(A、C、E、G)和呼吸速率。A、B:圣1号(兰德斯峡湾);C、D:浓度为0-2 mmol L-1的圣2号(Marselisborg Marina)和E,F:浓度为0-1 mmol L-1的Marselisborg Marina。G、H:St.3(汉斯霍姆)。比率平均为6个标准误差(n=3)。Michaelis-Menten动力学参数和Jassby和Platt动力学参数(括号内)均已给出。内政部:10.1371/期刊。波内。0105399.g006
在这项研究中,我们提出了一种测定浮游生物群落有氧呼吸速率的新方法,将超灵敏O2测定(在本例中是通过STOX传感器)与人工降低O2浓度的瓶培养相结合。通过该方法,氧气消耗率的分辨率与STOX传感器最佳工作范围内的氧气浓度成反比,即从饱和降至约10 nmol L-1 O2。假设14小时间隔内的信号漂移小于2%(图2),在100 nmol L-1 O2下可获得约0.4 nmol L-1 h-1的呼吸速率检测限,并且在500 nmol L-1 O2下可获得约2 nmol L-1 h-1的分辨率。这应与温克勒滴定法进行比较,温克勒滴定法的初始值和最终值之差的准确度最多为0.15 mmol L-1[2]。对于14小时的培养,这对应于约10 nmol L-1 h-1的检测限。培养时间的延长降低了Winkler方法的检测限,比本文介绍的基于STOX的方法要低得多,但我们发现,如果培养时间超过20小时,呼吸速率会增加数倍[22]。因此,与之前的直接测量相比,基于STOX的方法将O2呼吸测量的灵敏度提高了一个数量级以上。因此,它还允许测定深海水域、贫营养区和缺氧区的极低速率,低至约1 nmol-1 h-1[33]。对于居住在墨西哥海岸附近ETNP OMZ中间深度的浮游生物群落,在14小时长的瓶培养过程中测量了低于10 nmol L-1 h-1的CR率(图7)。
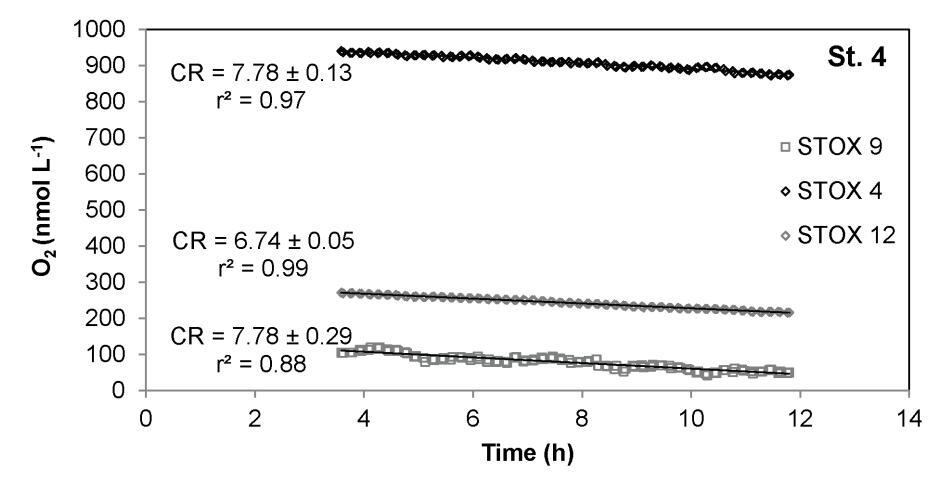
图7。与海洋海水孵育的STOX传感器中氧气消耗的时间过程。2012年4月,在海洋站4号进行的船上实验中,在3个重复瓶中氧气消耗的时间过程。在3种不同DO水平下孵育的3个样品的CR率相似(nmol-1 h-1)。CR率6标准误差通过线性回归确定,实验的前三个小时由于传感器信号上的噪声而被丢弃。内政部:10.1371/期刊。波内。0105399.g007
用这种方法测得的速率与传统温克勒滴定法测得的速率之间有很好的一致性(图6),证实了这种新方法的有效性。此外,该协议表明,通过在短时间间隔内人为降低样品中的氧浓度,对CR率缺乏可检测的直接影响;只要样品暴露在明显高于社区Km值的环境中。此外,该方法具有高分辨率和连续测量耗氧量的特点,为研究自然样品中的动力学参数以及浮游生物群落对不同O2水平的响应提供了独特的可能性。
丹麦三个测站(图S1)(0.16–0.8 mmol L-1 O2 h-1)测得的呼吸速率与先前对公海上层混合层的估计或测量值处于相同范围[1]。研究发现,调查沿海地区的地表水(,10 m)的平均呼吸速率为0.3 mmol L-1 O2 h-1,相当于沿海海洋地区组合数据集(0.3 mmol L-1 O2 h-1)的先前平均值[2,16,34]。这些监测站表现出不同程度的活动,正如人们所预期的那样,这些地点受到不同程度的富营养化和有机负荷的影响。St.1位于一个长而浅的河口,受两条有机负荷较高的河流排放的淡水的影响[21],其吸收率最高,并且可能有一个非常活跃的浮游生物群落。表现出较低呼吸率的St.2位于奥胡斯港外的沿海卡特加特海水中,有机负荷较低,浮游生物群落密度较低;St.3位于北海相对不太活跃的水域。最后,St.4是在开阔海域发现的低流速的一个例子。St.4处仍然相对较高的耗氧量可归因于取样深度(110 m)处存在深层次叶绿素最大值,同时细胞密度较高(参见Ulloa等人2012[35],了解次叶绿素最大值的一般描述)。此外,呼吸速率的差异也反映了浮游生物活动的季节性,夏季较高,秋季和冬季较低,与chl a浓度的季节性波动趋势相似(表1)。
然而,本次调查的目的不是为了确定丹麦沿海水域的准确呼吸速率,而是为了调查使用人工降低O2浓度的水测定呼吸速率的方法的有效性,以及测定低O2浓度下海洋和河口水中O2吸收的动力学。因此,我们没有试图密切模拟现场温度条件,也没有研究呼吸速率随昼间光周期的变化,因为光周期刚过,呼吸通常较高[36]。我们决定通过直接建模氧气浓度随时间的变化来估计氧气吸收的动力学参数,以避免线性化过程[37]的不准确和限制,以及使用因变量和推断值(即斜率)代替测量值的连续误差。Michaelis-Menten模型似乎对所调查的混合浮游生物群落的动力学进行了合理的描述,尽管该方程并不严格适用于具有不同动力学参数的异质种群[38]。
通过应用米氏动力学,我们假设聚集体中的扩散限制可以忽略。事实上,当细菌聚集时,由于邻近生物体的消耗,它们会经历较低的外部O2浓度。在厚生物膜的极端情况下,O2的吸收可以用半级动力学来描述[39]。浮游生物群落中总会有一些聚集,这将导致明显的Km值高于细胞值。随着电池尺寸的增加,扩散到电池的效率降低[40],因此,我们可以预期大型电池的表观Km值会更高。然而,海水中群落呼吸的主要部分(.70%)可归因于尺寸等级为10 mm的生物体,而尺寸等级小于1 mm的生物体高达58%,通常以细菌为主[41–44]。由于未完全满足Michaelis-Menten模型的假设,使用公式2建模的曲线与实验数据非常吻合,但导致建模的Vmax与实验检测到的最大呼吸值不匹配。因此,我们尝试应用Jassby和Platt(1976)的经验模型,在这种情况下,我们发现了更好的一致性。两种模型之间的主要区别在于,对于相同的假设Km值,通过改进的Jassy和Platt模型,在较低浓度下接近最大速率(Vmax)(图S3)。开发原始模型是为了更好地模拟混合微生物群落的行为,如本研究中所研究的微生物群落。
在我们的反应堆实验中估算的表观Km值低于之前报告的海洋细菌[45]。然而,在这个数量级上,O2的半饱和常数在微生物界是一个广泛存在的现象[46–49],以及通过高亲和力末端氧化酶从极低O2浓度下的有氧呼吸中获取能量的能力[49–51]。事实上,O2的还原是一种(火用)反应,它可以提供足够的自由能,以在极低O2浓度下为细胞过程提供燃料[50]。因此,毫不奇怪,只要氧气可用,微生物就可以使用氧气作为电子受体,即使浓度只有几纳摩尔[52]。
对来自公海和沿海地区的海洋样品进行的元基因组分析表明,Km值为几纳摩尔的高亲和力末端氧化酶的丰度非常低,而对培养细菌的分析表明,在734个受调查需氧菌中,93%存在此类氧化酶[49]。海水中缺乏这种高亲和力末端氧化酶并不令人惊讶,因为大部分海洋都是完全有毒的,只有动物内脏中的细菌[53],大型粪便颗粒和海洋聚集体中的细菌[54–56]暴露在高度降低的环境O2浓度下,它们可能会经历纳摩尔范围内的O2浓度。另一方面,Kalvelage等人最近在含氧量最低的水域中报道了高亲和力末端氧化酶[57]。然而,低亲和力末端氧化酶的Km值约为200 nmol L-1,因此即使没有任何高亲和力末端氧化酶的浮游生物群落也可能具有如此低的Km值。我们的一些数据也表明,细菌群落更为多样的沿海环境和低氧适应群落的海洋最低含氧量区的特征是明显的Km值略低于200 nmol L-1。
呼吸速率可能与水的预处理有关。这可以从第二次耗氧实验(图5和表1:St.2*,2011年9月,6–14小时)中对Km值的不同估计中看出。长期暴露于完全缺氧是应该避免的一个因素,因为它通常会导致非常不同的速率和Km值。此外,由于应允许社区适应新的条件,应丢弃孵化前1-2小时的数据[58]。建议使用2至16小时的数据确定动力学参数。超过16–20小时的孵化可能会导致高度错误的数据,至少在公海水样中是如此(L.Tiano等人在准备中)[22]。
我们意识到微生物过程体外测量的局限性[15,16,59],但在所介绍的装置中使用STOX传感器为解决与先前测定海水呼吸速率相关的几个问题提供了一个极好的解决方案。事实上,它是一种实时评估浮游呼吸的简单而准确的方法,不需要使用转换因子和估计值。例如,电子传输系统(ETS)方法是一种间接方法,必须通过经验确定的算法将测量的潜在呼吸率转换为体内呼吸率[60]。另一种间接方法是根据颗粒有机物(POM)通量进行估算[33],这种估算受到许多偏差的影响[5,61]。
STOX传感器的高灵敏度允许非常短的培养时间。此外,在整个实验过程中,反应器中O2的减少在浓度超过估计Km值三倍时呈线性,这意味着稳定状态社区[16]。因此,我们排除了通常与长期孵化相关的社区组成的重大变化[62]。
电化学传感器已应用于本研究中,但也有可能使用新一代光电二极管。这些新开发的非常灵敏的光电二极管是STOX传感器的一种有价值的替代品[20]。无论是否使用电化学传感器或光电二极管,都应在记录氧气消耗前后进行仔细的现场校准,以验证传感器是否存在污垢。事实上,聚集体可能在STOX传感器的尖端周围形成;这种现象在富含硅藻的沿海水域(L.Tiano,个人观察)中观察到,光电极表面可能形成生物膜,从而导致错误的测量。
通过使用本文所述的技术,可以比较O2吸收速率与其他微生物代谢过程的速率,如通过高分辨率15N同位素技术获得的N-转化速率,并研究从有氧呼吸到无氧呼吸的转换[63,64]。我们现在还可以详细研究定义的低O2浓度如何影响自然环境(如海水(T.Dalsgaard,个人交流)或纯培养物中的基因表达[22]。
图S1采样点地图。STOX培养方法通过来自丹麦沿海和峡湾水域3个站点的样本进行测试(左侧地图)。丹麦采样地点(N):1号站、兰德斯峡湾、2号站、马塞利斯堡码头、3号站、汉斯霍姆。所有样本均在距海岸线100 m的范围内采集。该方法还用于评估热带北太平洋东部(ETNP)氧气最低区(右侧地图)水样中的浮游生物群落呼吸速率。4号站,距离墨西哥海岸700公里。该地图显示了代表150 m深度处以饱和度百分比表示的氧浓度的等高线,氧数据来自2009年世界海洋地图集,美国国家海洋数据中心(海洋数据视图)。(TIFF)
图S2孵化装置示意图。监测氧气消耗的设置,包括:1)换档箱。2)前护罩的电源。3)ACD816bit A/D转换器(Unisense A/S)、4)PA8000多通道皮安计(Unisense A/S)、5)计算机、6)磁搅拌器(IKA、实验室光盘)上的水浴,其中包含装有STOX传感器和玻璃涂层磁铁的玻璃瓶。在大约一米的实验台空间上,可以安装实验所需的所有设备,最多8个复制瓶和8个STOX传感器。(TIFF)
图S3动力学模型的比较。上图:呼吸速率与氧气浓度,使用相同固定参数比较Michaelis-Menten和Jassby及Platt方程。下图:第2站一个样本孵化过程中氧浓度的时间演变,以及比较Michaelis-Menten、Jassby和Platt方程,使用100%氧饱和度(173 nmol L-1 h-1)下测得的呼吸速率作为Vmax。(TIFF)
文本S1 STOX传感器的耗氧量。(DOC)
文本S2不同动力学模型使用的比较和评估。(DOC)
 相关新闻
相关新闻