热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
许多物理和化学过程控制异化 Fe(III) 还原细菌对 Fe(III) 羟基氧化物的还原程度。 次生铁矿物在 Fe(III) 羟基氧化物上的表面沉淀限制了微生物 Fe(III) 还原的程度,但这种现象尚未在自然界中观察到。 本文报告了在深达 10 厘米的淡水沉积物中水铁矿表面的次生铁矿物(针铁矿)结壳的观察结果。 沉积物表面的特点是水铁矿占优势,具有生物茎和鞘。 通过 16S rRNA 基因分析在 1 和 2 厘米的沉积物深度检测到 Fe(II) 氧化细菌(Gallionellaceae)。 沉积物孔隙水中的 Fe2+ 浓度在 2-4 cm 深度相对较高。 在 1、2 和 4 cm 深度检测到与异化 Fe(III) 还原菌相关的 16S rRNA 基因。 Fe K 边缘扩展 X 射线吸收精细结构 (EXAFS) 分析的结果表明在 3 cm 以下深度存在针铁矿和菱铁矿。 然而,尽管在 4 厘米以下的深度存在丰富的水铁矿,但铁矿物成分的变化仅限于 3 到 4 厘米之间的沉积物深度。 在深度超过 6 cm 处观察到高浓度增加。 孔隙水中生物质的稳定同位素分析表明,7 cm以下深度发生了乙酰碎屑岩生化。 透射电子显微镜观察表明,在深度低于 3 cm 的茎和鞘上存在针铁矿和菱铁矿。 转换电子产额 EXAFS 分析结果表明针铁矿在 10 cm 深度占主导地位,从而表明水铁矿在该深度被针铁矿包裹。 此外,下面为4cm深处的水铁矿的不完全降低不是由于缺乏有机碳的,但可能是由于针铁矿对水铁矿的表面结垢。
Fe(III) 羟基氧化物的微生物还原是缺氧沉积物环境中 Fe 和碳循环的关键过程 (Lovley, 1991; Nealson & Saffarini, 1994)。 将 Fe(III) 还原与有机碳或 H2 的氧化结合起来的异化 Fe(III) 还原菌,在自然环境中,尤其是在非硫化还原条件下,在有机碳的氧化中起重要作用 (Lovley, 1991 ;洛夫利,1994 年)。 已从天然样品中分离出各种异化 Fe(III) 还原菌,它们能够诱导各种 Fe(III) 羟基氧化物的还原溶解与有机碳的氧化相结合(Lovley 和 Phillips,1988 年;Lovley 等人,1993 年) ;Coates 等人,1996 年;Slobodkin 等人,1997 年)。 例如,自然环境中最常见的异化 Fe(III) 还原之一将乙酸盐氧化与短程有序 Fe(III) 羟基氧化物的还原结合起来,可以用以下方程表示 (Roden, 2003):
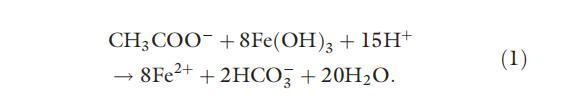
由于异化 Fe(III) 还原菌和其他微生物(氧化根还原剂或产甲烷菌)对同一电子供体的竞争,异化 Fe(III) 还原抑制微生物的氧化根还原和还原生产 (Lovley & Phillips, 1986a , 1987a; Chapelle & Lovley, 1992; Roden & Wetzel, 2003)。 乙酸盐不仅可以被异化 Fe(III)-还原菌利用,如方程式 (1) 所示,也可以被氧化根还原剂和产甲烷菌利用,如下所示 (Lovley & Klug, 1986):
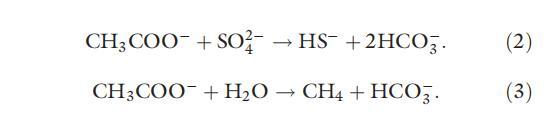
异化 Fe(III) 还原菌可以将醋酸盐和 H2 的浓度维持在过低的水平,使硫酸根还原剂和产甲烷菌无法发挥活性(Lovley & Phillips, 1987a; Roden & Wetzel, 2003)。 因此,Fe(III)羟基氧化物对异化Fe(III)还原菌的抑制作用与微生物氧化根还原和可利用性产生密切相关。
Fe(III) 羟基氧化物被异化 Fe(III) 还原菌还原的程度由各种物理和化学因素决定。 控制 Fe(III) 羟基氧化物还原程度的一个重要因素是 Fe(III) 羟基氧化物的矿物学。 该领域的开创性研究表明,短程有序的 Fe(III) 羟基氧化物,例如水铁矿,比结晶 Fe(III) 羟基氧化物更容易被异化 Fe(III) 还原菌还原 (Lovley & Phillips, 1986a,b ;洛夫利,1991 年;菲利普斯等人,1993 年)。 最近的研究表明,Fe(III) 羟基氧化物的还原程度与矿物中结构紊乱的数量和/或比表面积呈正相关,而与存在的 Fe(III) 矿物种类无关(Roden 和 Zachara, 1996;Zachara 等人,1998 年;Roden,2003 年;Hansel 等人,2004 年)。 在地下水排放点、热液喷口和湿地等自然环境中形成的 Fe(III) 羟基氧化物通常是短程有序的 Fe(III) 羟基氧化物,例如水铁矿 (Emerson & Revsbech, 1994; Banfield et al., 2000;肯尼迪等人,2003 年;Gault 等人,2011 年)。 此外,由于涉及各种生物有机碳(例如,鞘、茎和细胞外多糖)和无机离子(例如,硅酸盐、铝和磷酸盐),观察到结构上比合成水铁矿更不有序的 Fe(III) 羟基氧化物在天然沉积物中(Toner 等人,2009 年,2012 年;Edwards 等人,2011 年;Mitsunobu 等人,2012 年)。 天然 Fe(III) 羟基氧化物与异化 Fe(III) 还原菌(例如,Geobacter metallireducens 和 Shewanella putrefaciens)的孵育表明,Fe (III) 羟基氧化物的微生物还原是快速且大部分是完全的; 因此,天然 Fe(III) 羟基氧化物对于异化 Fe(III) 还原菌的可用性很高(Straub 等人,1998 年;Zachara 等人,1998 年;Emerson,2009 年;Langley 等人,2009 年)。
控制微生物 Fe(III) 还原程度的另一个重要因素是 Fe(III) 羟基氧化物对异化 Fe(III) 还原细菌的可及性。 如方程 (1) 所示,异化 Fe(III) 还原生成 Fe2+ 离子,其中一部分沉淀为次生 Fe 矿物,如针铁矿、磁铁矿、菱铁矿、紫铁矿和绿锈(Lovley 等,1987b; Fredrickson 等,1998;Liu 等,2001;Benner 等,2002;Zachara 等,2002;Hansel 等,2003、2004、2005;Pallud 等,2010a、b)。 随后,残留在水中的部分 Fe2+ 离子可能会吸附在矿物表面(Roden & Zachara,1996;Urrutia 等,1998;Liu 等,2001)。 次生 Fe 矿物的沉淀和 Fe(III) 羟基氧化物上 Fe2+ 离子的吸附已被建议降低 Fe(III) 羟基氧化物还原的程度(Roden 和 Zachara,1996 年;Urrutia 等人,1998 年;Roden 和 Urrutia, 1999, 2002; Roden et al., 2000; Pallud et al., 2010a,b),可能是因为这些反应限制了 Fe(III) 羟基氧化物对细菌的可及性。 因此,即使自然环境中的短程有序 Fe(III) 羟基氧化物在其矿物学方面具有生物可利用性,但由于次生 Fe 矿物的过度结壳和/或 Fe2+ 吸附引起的有限可及性可能会限制或阻止天然 Fe(III) 羟基氧化物。 此外,即使在短程有序的 Fe (III) 羟基氧化物的存在下,如果它们既不可接近也不可用,则根硫酸还原剂和产甲烷菌对有机碳的氧化可能不会受到抑制。 汉塞尔等人。 (2003) 通过实验室实验观察到,当针铁矿在水铁矿表面沉淀时,会发生水铁矿的不完全还原。 然而,只有少数研究调查了在野外条件下次生铁矿物的表面结壳与 Fe(III) 羟基氧化物还原程度之间的关系。 此外,在实验室或实地研究中尚未对 Fe(III) 羟基氧化物上的次生铁矿物结壳进行定量表征。
在本研究中,我们研究了次生 Fe 矿物对 Fe(III) 羟基氧化物的表面结壳,这可以限制微生物 Fe(III) 羟基氧化物在该领域的还原程度。 本研究调查了从日本广岛大学的 Budo Pond 收集的淡水沉积物。 Takahashi 等人先前已研究过该区域。 (2007) 和加藤等人。 (2012),他报道了具有生物茎和鞘的 Fe(III) 羟基氧化物存在于该领域。 以下是我们期望:(ⅰ)的Fe(III)羟基氧化物存在于淡水沉积物是短程有序,而铁(III)羟基氧化物的一部分被转换成次级铁矿物质由异化Fe(III)还原,并(ii)通过二次铁矿物的Fe(III)羟基氧化物的结壳降低的Fe(III)羟基氧化物的可接近性,这反过来又增加在水中的甲烷生成活性,尽管短rangeordered的Fe(III)的存在下羟基氧化物沉积物中. 为了定量检查次生铁矿物表面结壳的存在,应用了两种扩展 X 射线吸收精细结构 (EXAFS) 模式,如下所示:(i) 传输模式,提供大量铁矿物信息,以及 (ii)转换电子产率 (CEY) 模式,可提供有关在颗粒表面形成的 Fe 矿物的信息。 我们使用微电极分析、透射电子显微镜 (TEM)、对沉积物的稳定同位素分析和 16S rRNA 基因分析对沉积物进行物理化学和微生物学表征。 本研究的结果将为进一步研究二次铁矿物结壳在控制微生物还原 Fe (III) 羟基氧化物方面的重要性提供基础。
 相关新闻
相关新闻