热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
物理和生物背景数据
11月,水柱与 -1.5和-1.7之间的温度和盐度 大约 34.6–34.8%。 荧光为零 从表面到底部,Calanus 种群集中在 100 m 以下的深度(结果未显示)。 7 月,水体分层有新鲜 (29.2 %) 和温暖的 (+7.2 冷却) 上层 5-10 米表层。 从 10 m 下降到 50 m 深度,温度下降 急剧增加到-1.2℃,盐度增加到34.6%。 荧光在 5 m 处达到峰值,对应于 1.14 lg chl a l-1 . 在 50 m 以下,藻类生物量非常低 (\0.08 lg chl a l-1 )。 卡兰努斯种群主要是 在上部 100 m 中发现(结果未显示)。
代谢
C. glacialis 在四种不同环境中的 C 特定需求 处理方法如图 2 所示。 问题,11 月的 Dark+FSW 治疗第 4-9 天没有结果。 第 1 天的结果 因此,将 3 至 3 用作第 4-6 天和第 11 月 7-9 日。
表 1 每个时间间隔的 C 特定需求率的双向 ANOVA,包括食物处理 (Food/FSW) 和光照状态 (Light/FSW) 暗)为主要因素
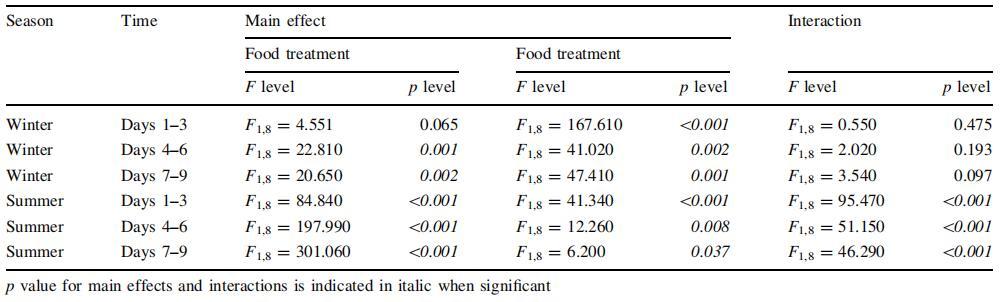
11 月,只有光线对 前 3 天的 C 特定需求(双向方差分析, 对于光,p\0.05,表 1)。 在那段时间里, 桡足类通过将其 C 特定需求增加 3 倍(单向方差分析,p\0.05),在存在和不存在 食物(表 1;图 1)。 在第 3-6 天和第 7-9 天, 光和食物对 C 特定需求有显着影响, 但不是关于它们的相互作用(双向方差分析,p\0.05 对于两个单独的因素,表 1)。 C 特定需求 在有食物的情况下保持高位,在结束时达到顶峰 实验结果为 3.8 倍 基线(单向方差分析,图 2)。 在缺少...之下 食物,C 特定需求在 3 天后下降, 虽然从未达到初始低基线值 测量 Dark+FSW(图 2)。 在没有光线的情况下 但食物的存在(黑暗+食物处理),桡足类 C 特定需求在 3 天后增加了约 1.3,并且 仍然显着高于基线(图 2) 在整个实验过程中,虽然从未接近 在治疗中测量的相对较高的值 光+食物。
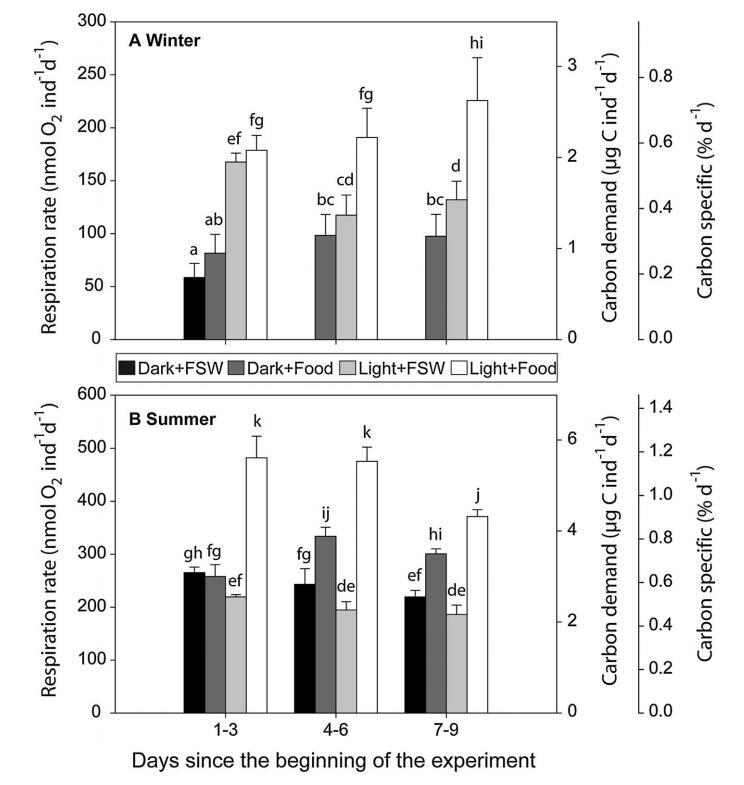
图 2 呼吸速率,碳 每个人的需求率, 和碳特定需求 天 (nmol O2 ind-1 day-1, lg C ind-1 day-1 和 % day-1 分别)和标准 每次处理的偏差, 黑暗和过滤的海水 (黑暗+FSW),黑暗和食物 加成(暗+食物),光 暴露和过滤海水 (Light+FSW) 和 Light and 食物添加 (Light+Food),对于 一个冬天和一个夏天。 因为 技术问题,没有结果 可用于第 4-9 天 Dark+FSW 中的治疗 十一月。 对于所有碳特定需求结果, 不同的字母表示 之间的显着差异 处理(单向方差分析, 迷幻剂; p \0.05)。 请注意 每个不同的 y 轴比例 季节
7 月(图 2b),基准 C 特定需求 (黑暗 + FSW;第 1-3 天)是 *3 高的因子(t 检验 p\0.05) 比 11 月。 这是综合效果 光和食物的两个因素(双向方差分析) 显着影响了夏季的特定碳需求, 尽管随着时间的推移,食物似乎比光更重要。 事实上,3天后,用食物治疗的生物体, 无论光照条件如何,都具有更高的 C 特异性 与无食物处理相比,需求增加了 1.25-1.8 倍(单向方差分析,图 2)。 这 没有食物的治疗中的 C 特定需求保持不变 低 (Light+FSW) 甚至随着时间的推移而降低 (Dark+Food,单向方差分析,图 2)。
讨论 食物的供应被认为是最重要的 浮游动物呼吸和碳需求的因素 (高桥等人,2002 年)。 已发现喂食过量食物的 C. hyperboreus 会增加其呼吸速率 系数 3(Takahashi 等人,2002 年),相反, 缺乏食物可能会减少桡足类动物的呼吸和 排泄率(Ikeda 和 Skjoldal 1989;Ikeda 等人。 2001)。 在本研究中,暴露在光线下也是 考虑到,光和食物都被发现 影响冬季和夏季的桡足类 C 特定需求。
C. glacialis CV 的碳需求范围为 0.72 lg C ind-1 day-1 in 11 月 (Dark+FSW) 到 5.52 lg C ind-1 day-1 in 7 in 7 当同时暴露于两者时 清淡+食物(图2)。 波弗特海东南部的 C. glacialis 雌性也对 生物生产季节,但他们的呼吸是 远高于本研究中测量的 CV,增加 从 4.8 到 7.2 lg C ind-1 day-1 三月到 5 月 14.4–21.6 lg C ind-1 day-1(Seuthe 等人,2007 年)。 这些对女性更高的碳需求可能是 解释为女性较高的代谢活动由于 性腺成熟和产蛋期间的投资 冬春过渡期(Hirche 和 Kattner 1993)。 11月,原位碳需求非常 C. glacialis CV 较低,表明这些桡足类 处于滞育状态。 尽管如此,他们很快就做出了回应 光和食物通过增加呼吸作用 3.5 对应于原位碳需求量测 对于 7 月的活跃 C. glacialis CV。
光似乎是唤醒滞育桡足类动物的主要诱因,但如果没有食物,碳 需求再次降低。 碳需求的事实 实验从未恢复到低的原位值 表明他们要么需要更多时间来减少 新陈代谢或恢复滞育是不可能的 一旦他们被唤醒。 后者意味着 唤醒桡足类动物将不得不应对更高的碳 直到有食物供应。
结果表明,在冬天,当桡足类 滞育,光是一个主要的“唤醒”因素。 这可能是由于光是初级生产的限制因素,因此是桡足类动物的藻类食物。 冬天。 从充足的光线开始大约需要 2-3 周 可用于初级生产,直至达到藻类生长高峰期 达到(Engelsen 等人,2002 年;Leu 等人,2010 年)。 这个 时间滞后类似于激活所需的最短时间 越冬Calanus中的消化酶(Hirche 1983)。 桡足类光的剧烈变化研究 代谢可能与大多数温带或 热带地区,但对于极地地区,这是至关重要的 重要性。 在北极,海冰破裂的时间 在同一地区,每年可能会有很大差异 (http://polarview.met.no/)。 较早的光线穿透 由于海冰条件变化而导致的水柱 (Arrigo et al. 2008) 很可能会触发桡足类动物 “醒来”,从而为即将到来的事情做好准备 藻类食物。 对光等外部线索做出反应,即使 在处于某种休眠状态时,将因此确保 C. glacialis 来“醒来”并及时为浮游植物盛开做好准备,这是一年中的主要食物事件。
在夏天,与冬天相反,食物似乎是一种 桡足类碳需求增加的主要因素。 这 初始碳需求基准率测量 7 月的黑暗 + FSW 与测量的比率相似 Bamstedt 和 Tande (1985) 的 C. glacialis CV 类似 西巴伦支海 6 月的低温。 这些值低于先前报告的碳 C. glacialis 雌性在类似条件下的需求 (Bamstedt 和 Tande 1985;Ikeda 和 Skjoldal 1989; 苏特等人。 2007)。 这可能是由于较大的尺寸 与 CV 相比,女性的数量和增加的碳 对繁殖雌性的需求(Hirche 和 Kattner 1993)。 不管光照条件如何,饥饿的桡足类动物 在为期 9 天的实验中降低了他们的新陈代谢 七月。 相反,在有食物的情况下,碳需求 保持高位。 这表明滞育可能是 由缺乏食物触发,即使光照仍然存在。 然而,储存的脂质量可能也起到了作用 何时下降到深度和减少的重要作用 代谢(Miller 等人,2000 年;Søreide 等人,2010 年)。 在 2010年7月,采样点藻类生物量偏低 C. glacialis CV 中的脂囊大小相当大(JE 瑟雷德人。 观察。)表明这些桡足类动物很快 准备越冬。 从 8 月中旬开始,C. glacialis CIV 和 CV 主要出现在 Billefjorden 的深处(JE Søreide pers. observ.)。
 相关新闻
相关新闻