热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
结果
3.1 、 海藻在为期六天的实验室实验中适应升高的 pCO2
物种对升高的 pCO2 的反应不同(物种 * pCO2 相互作用,F10,55 = 20.51,p < .001,另请参见图 1)。 在 6 天的过程中,轻度升高的 pCO2 水平会导致所有物种的生长增加。 然而,pCO2 水平超过 700 latm 会导致所有非开花形成物种的生长减少,但在开花形成物种中不会,其中 pCO2 水平增加的反应规范是指数级的(策略 * pCO2 相互作用,F1,12 = 9.06,p < .001,另请参见图 1)。 最好地描述这些数据的模型包括物种或嵌套在策略中的物种,但不包括“策略”本身作为一个因素。 在非开花形成物种中,生长对升高的 pCO2 的响应遵循更单峰的分布,在 700 latm pCO2 附近获得最大生长。
3.2 、 海藻在为期一个月的室内中观实验中适应升高的 pCO2
在室内中层环境的 28 天适应期内,我们发现大型藻类物种的反应方式与实验室实验中发现的相似。 物种之间存在显着差异(物种 * pCO2,F10,44 = 7.36,p < .0001,参见图 2,以及图 S8 和 S9),形成水花的物种和未形成水花的物种的反应方式存在明显差异到升高的 pCO2(策略 * pCO2,F2,30 = 64.84,p < .0001,见图 2)。 在这里,我们发现对嵌套在“策略”中的“物种”的支持比单独的“物种”更有力,解释了对 pCO2 升高的反应。
在 400–2,000 latm pCO2 的范围内,形成水华的石莼物种的生长速率随着 pCO2 的升高而增加,尽管与实验室实验不同,在较高的 pCO2 水平下,这种增加变得不那么明显(比较生长从 400 增加到 1,000 latm:1.6 倍和 1,500 至 2,000 latm:1.01 倍)。 外推的最大增长率在大约 3,000 latm pCO2 时达到(图 2),而非开花形成物种显示出单峰响应曲线,最高增长率平均达到 724 ? 41 latm CO2(图 2),这与实验室设置中这些物种的生长开始下降的 pCO2 水平相似(另请参见图 S7)。
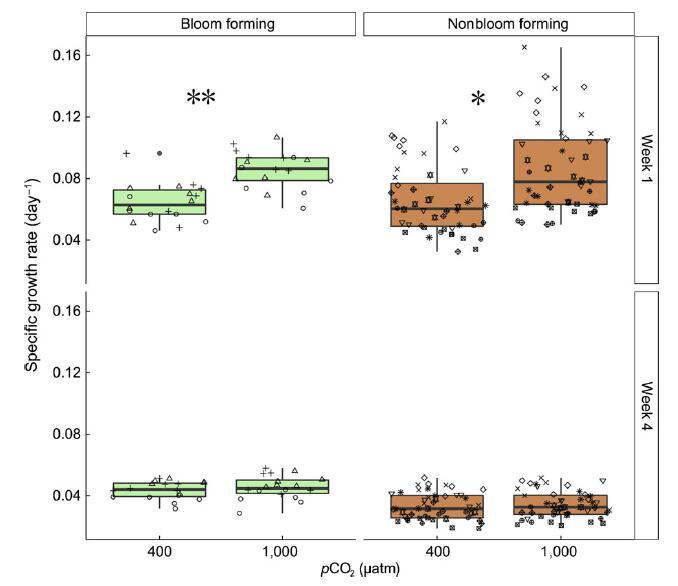
图 3 特定增长率 成花(淡绿色)和非成花 (浅棕色)大型藻类在 环境 (400 latm) 和高架 (1,000 latm) 室外 pCO2 水平 中观表现出显着差异 在如何形成绽放和 非开花物种响应 半自然环境中的碳富集 设置(F2,9 = 5.0,p < .05)。 在这两种情况下, pCO2 升高对 增长率(* 表示 p < .05,** 表示 p < .01) 在第一周 实验。 然而,在上周 (第 4 周),增长率下降,并且 没有明显区别 pCO2 处理之间。 箱线图 按标准显示,每个 n = 6 物种和 pCO2 水平和形状 指出个别物种,见 图 2 为图例。 【彩图可以 在 wileyonlinelibrary.com 上查看]
然而,在所有处理和分类群中,第 1 周(另请参见图 S10 和 S11)后,生长率急剧下降,并持续到实验结束,除了对照池中的石莼,可能至少部分是在反应中适应水箱设置本身,但很可能主要是由同时发生的温度升高驱动的。 虽然不一定是因果关系,但随着时间的推移温度升高与增长率下降之间存在很强的相关性,尤其是在较高的 pCO2(线性模型 Δ SGR 随时间变化 ~ Δ 温度(°C)随时间变化,r2 = . 38,p < .001,混合模型 pCO2 * 温度 * 策略,F4,40 = 3.068,p < .001,包含温度的模型的 AICc 分数低于不包含温度的模型,表明 pCO2 的变化仍然是响应)。
详细地说,lmax 平均下降了原始 lmax 的 0.33 倍(“周”的影响,F2,8 = 15.56,p < .0001)。 在较高的 pCO2 条件下 4 周后,所有形成水华的物种仍保持较高的生长速率(与 400 latm 的第一周相比)(图 2 下图)。 在非开花形成物种中,4 周后的生长率在所有处理中均低于第一周(事后 p < .05)。
此外,非开花类群响应的变化也随着时间的推移而减少(图 2,比较顶部和底部面板上的物种分布),以至于同一“策略”的单个物种之间的差异不再显着不同。
3.3 、 海藻在为期一个月的沿海中观实验中适应升高的 pCO2
在沿海中宇宙中,建立了两个 pCO2 水平,即 400(环境)和 1,000 latm(IPCC 情景 RCP 8.5 下的 2,100 预测)。 第一周后的反应(图 3 顶部面板,星号表示显着性)与室内中宇宙中相同 pCO2 水平的反应广泛一致(见图 4),其中在石莼花形成物种中的反应最强(生长倍数变化:1.33),以及非开花形成物种的显着但不那么强烈的反应(非开花形成物种的平均倍数变化:1.25,策略 * pCO2,F2,9 = 5.0,p < .05)。 在室外中观中,对于任一策略(pCO2 * 位置 * 策略,F1,6 = 3.12,p = .13)的物种,生长率的初始响应与室内设施中的初始响应无法区分。 与室内中层一样,所有处理的增长率在实验期间均下降(图 3 下图,再次强烈表明温度升高会产生影响,F3,50 = 10.458,p < .0001,另请参见图 S12)。 对升高的 pCO2 响应的变化在室外中层中通常比在室内中层中大(见图 S13)。
3.4 、光合作用和呼吸速率的变化响应于室内中层环境中 pCO2 升高
对在室内中层环境设施中生长的样品测量的净光合作用(NP,第 1 周的图 4a,所有周的图 S14)和呼吸(RE,第 1 周的图 4b,所有周的图 S15)显示出与生长相似的响应对升高的 pCO2 有显着不同的反应(NP:物种 * pCO2,F4,10 = 63.99,p < .0001,RE:F4,10 = 28.57,p < .0001,图 4)。 石莼物种的 NP 和 RE 随 pCO2 水平升高而增加,而其他物种则表现出大致单峰响应,峰值约为 700 latm pCO2。
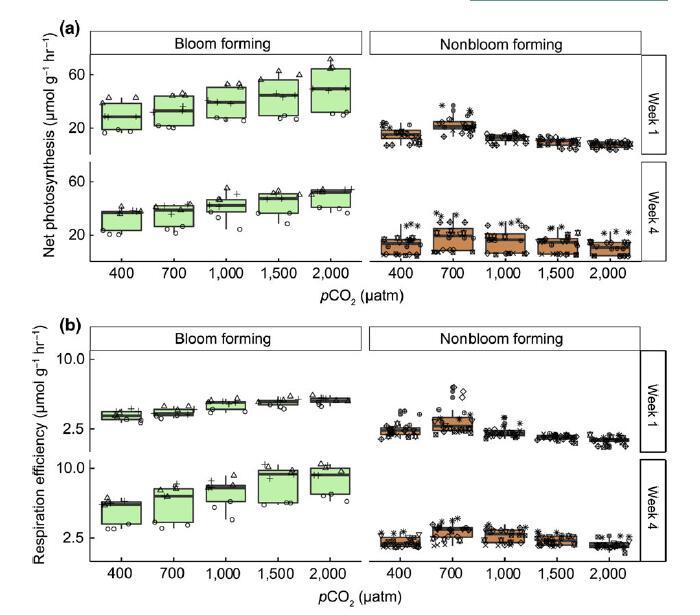
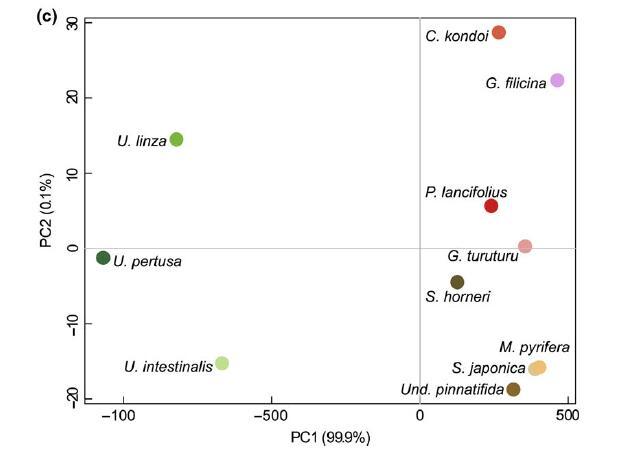
图 4 成花(淡绿色)和非成花的代谢性状(净光合作用和呼吸作用,lmol O2 g?1 鲜重 hr?1) (浅棕色)大型藻类对升高的 pCO2 水平的反应在反应范数的大小和形状上有所不同(a,NP:策略 * pCO2, F4,10 = 63.99, p < .0001, b, RE: 策略 * pCO2, F4,10 = 28.57, p < .0001)。 净光合作用(NP,a 组)和呼吸作用(RE,b 组) (有关光合作用与呼吸比率的信息,请参见 SI)受 pCO2、在中宇宙中花费的时间和海藻“策略”的影响 随着 pCO2 升高,形成海藻的速率总体上更显着增加,而代谢性状的反应规范在 不开花的海藻在很大程度上是单峰的,峰值速率约为 700 latm pCO2。 幅度明显下降 从第 1 周到第 4 周对光合作用和呼吸作用的反应,形成海藻的海藻的呼吸速率变化更大。 箱线图按标准显示,粗线表示中位数,胡须延伸到最高和最低值 在下/上四分位数的 1 个四分位距 (IQR) 内。 n = 6 每个物种每个 pCO2 水平。 形状表示单个物种,见 图 2 为图例。 基于 lmax、Km、呼吸和光合作用速率的主坐标分析 (PCA) (c) 绿色)和不开花(红色和棕色阴影)的物种在室内中层环境中。 开花和非开花物种分别聚集 彼此基于前两个主要成分(lmax 和光合作用速率)。 [彩色图可在 wileyonlinelibrary.com 查看]
将增长的变化与 NP 或 RE 的变化相关联(参见图 S17,r2 = .48,p < .001 和 r2 = .58,NP 和 RE 分别为 p < .001)进一步证实存在稳健性NP 或 RE 的变化与 pCO2 水平升高的生长变化之间的关系,即对生长有较大反应的物种,在代谢性状方面也表现出较大的反应。 与响应 pCO2 升高的增长率变化一样,我们发现在整个实验期间,NP 或 RE 对各组 pCO2 升高的反应强度降低(图 4a、b、pCO2 * 周:F3,30 = 24.69,p < .001),温度升高可以解释其中的一些原因(F4,40 = 13.10,p < .001)。 有趣的是,在整个实验中,光合作用与呼吸作用的比率(净光合作用/呼吸作用,图 S16)在形成水华的物种中基本保持稳定(例如,基本响应随时间几乎没有变化)。 然而,在非开花物种中,光合作用与呼吸比随时间的变化存在显着差异。 虽然第一周非开花形成物种的光合作用与呼吸比没有显着差异(事后 p = .13),但在培养 4 周后,出现了两种不同的策略(见图 S14 和 S15):G. filicina, G . turuturu、P. lancifolius、C. kondoi 在培养 4 周后都表现出相对于 RE 的 NP 升高的趋势,而在 S. japonica、U. pinnatifida 和 S. horneri 中,呼吸速率相对于光合作用速率增加。
在环境复杂性海洋环境中驯化的蓝藻和海藻能提高海洋中PCO2分压总保有量——摘要、介绍
在环境复杂性海洋环境中驯化的蓝藻和海藻能提高海洋中PCO2分压总保有量——材料和方法
在环境复杂性海洋环境中驯化的蓝藻和海藻能提高海洋中PCO2分压总保有量——结果
在环境复杂性海洋环境中驯化的蓝藻和海藻能提高海洋中PCO2分压总保有量——讨论、致谢!
 相关新闻
相关新闻