热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
水影响自然湿地和耕地,由于持续的气候变化,洪水的频率和强度在过去几十年有所增加。水稻作为一种作物,在雨水灌溉的低地也会受到洪水的影响,在这些地区,植物在生命周期早期面临很高的淹没风险。深水水稻的一个被充分研究的特征是其卓越的浮潜能力,其节间伸长高达30厘米d−1,使植物能够跟踪上升的水位。浮潜有助于从大气中获取氧气,从而维持内部通气。部分淹没开始后不久,茎被淹没部分的节点上生长出大量水生不定根。众所周知,在许多湿地植物物种中,茎节中出现不定根是对部分淹没的响应。这些根通常不会到达土壤表面,而是在洪水中漂浮,因此被称为水生不定根。深水水稻水中不定根的功能与从洪水中氮和磷的养分获取有关在本研究中,报道了两种类型的水生不定根的形成,在出现的时间和关键的形态学和解剖学特征上有显著差异。研究发现,叶鞘延迟了淹水后根系发育相关基因的时空表达,决定了根系出现的时间,甚至在淹水后期提高了根系的粗细和伸长能力。淹没是一种复合压力,因此,耐受性是由一系列特征赋予的。由于茎伸长和水中不定根的形成发生在淹没后不久,本研究假设了茎生长和不定根的调节是遗传相关的。研究验证了在深水水稻中促进茎生长响应淹没的qtl也促进不定根生长的假设。
Unisense微电极系统的应用
使用O2微传感器测量附着在茎节上的水生不定根的径向O2分布。从主茎上切下具8-10 cm长AR1或AR2节间的茎;AR1在这个位置有一些横向分支,在测量过程中这些分支仍然存在。带有一个目标根的茎块(额外的根被修剪掉)固定在一个金属网上,放置在一个小水族馆中,并浸入空气平衡的去离子水中。空气石也提供温和的气泡插入到水族箱中,以确保扩散边界层(DBL)之间恒定。使用氧气微传感器(OX-10;Unisense丹麦),从DBL内部的根表面内侧400-500μm开始,以10-25μm为步长进行径向测量。传感器的定位使用unisense微电极进行调整,水稻根部的视觉辅助使用一个臂架解剖徕卡显微镜。对pO2的分析一直持续到传感器尖端完全穿过根的石碑为止。在测量过程中,使用O2光电传感器(Unisense)和温度传感器监测大体积水的O2和温度。根据制造商的说明,每天使用两点校准校准两个O2传感器。
实验结果
赋予水稻抗洪能力的一个关键特征是生长不定根的能力,作为对淹没的反应。决定水生不定根(AAR)发育和特征的深水水稻遗传性状尚未得到评价。利用渐渗的近等基因系验证了深水水稻显著的茎伸长能力与数量性状位点1和12相关也促进了AAR的发展。深水水稻基因型NIL-12在茎节处有扩大区域,在那里有大量AAR作为对淹没的反应。在淹水3天(AR1)~7天(AR2)内形成了两种根系类型(AR1和AR2),出现时间明显,形态解剖特征差异较大。叶鞘提供的机械阻抗使AR2出现较晚,根系较粗,伸长能力较强,耐干燥能力较强。关键基因的上调提示在激活AAR中分生组织的共同贡献,增强了这些组织的发展,以应对淹没。形态和解剖特征表明,AR2比AR1更适应长期淹水。因此提出了AR2在深水水稻中的作用是一种进化防御策略,以应对周期性淹没。
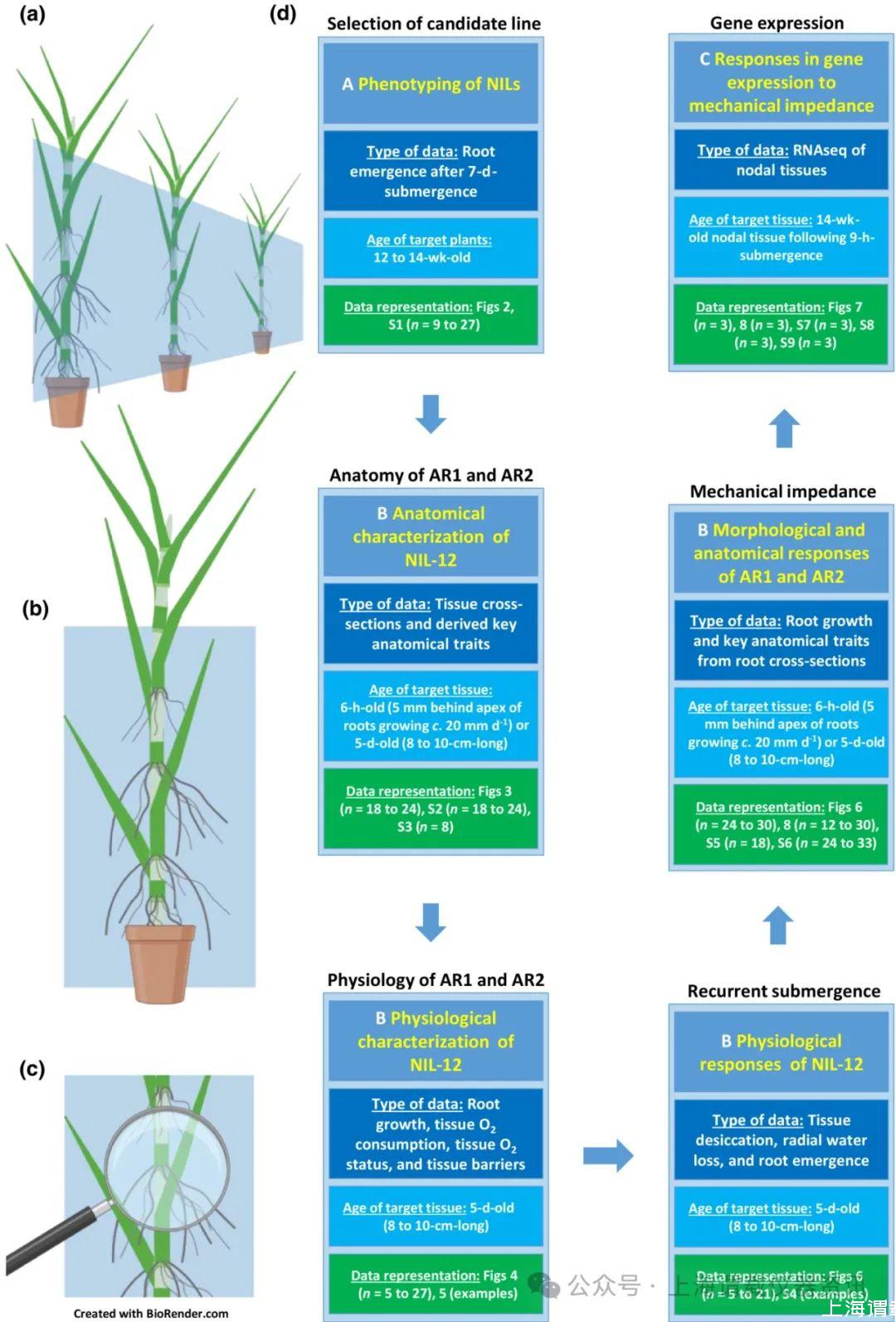
图1、展示实验条件和子实验相互关系的概念图。(a)不定根形成的表型作为对部分淹没的反应和目标基因型的选择。(b)nil12的解剖和生理特征。(c)基因表达对存在或不存在机械阻抗的响应。(d)数据类型、目标组织和数据表示。AR=不定根;NIL=近等基因系。
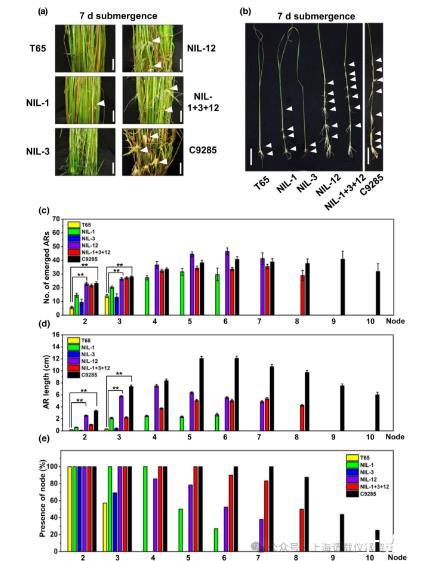
图2、深水水稻(Oryza sativa L.)QTL1和QTL12在水稻背景T65中促进不定根的形成。(a)T65、NIL-1、NIL-3、NIL-12、NIL-1+3+12和深水水稻C9285部分淹水7天。(b)去除叶片的代表性茎显示出不定根生长的节点(白色箭头)的位置。(c)T65、NIL-1、NIL-3、NIL-12、NIL-1+3+12和C9285浸水7天植株各节点(节点1为最年轻、最上面的节点)上出现的ar平均数量。(d)在指示节点上生长的ARs的平均长度由三个独立的实验确定。(e)T65、NIL-1、NIL-3、NIL-12、NIL-1+3+12和C9285浸水7天植株均有淋巴结。低地水稻T65有2-3个节,深水水稻C9285有10个节。AR=不定根;NIL=近等基因系。QTL=数量性状位点。
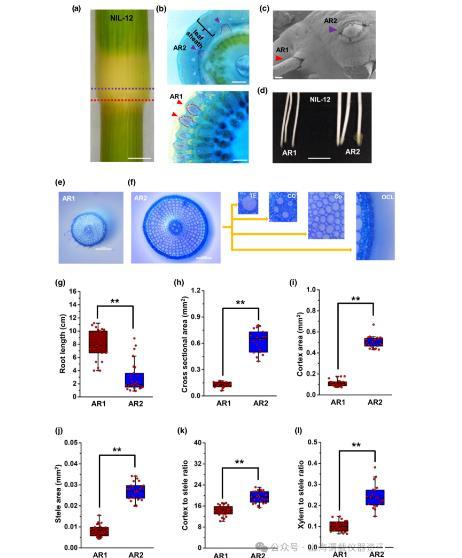
图3、水稻(Oryza sativa)的NIL-12株在其节点上产生两种不同的AR类型。(a)NIL-12茎节,用虚线紫色和红线表示不定根发展的两个位置。(b)(a)所示位置的节点截面。在较低位置发育的不定根称为不定根1,在被叶鞘覆盖的节点上部发育的不定根称为不定根2。(c)显示AR1和AR2的扫描电子显微照片。Bar,300μm。(d)有代表性的AR1和AR2伸长。棒,5毫米。(e)从NIL-12的第4年轻节的AR1和AR2的横切面在根尖后5mm切除,用甲苯胺蓝染色。(f)AR2显示气管元件(TE)、中心柱体(CC)、皮层(Co)和外细胞层(OCL)。横切面取根尖后5mm。棒材,0.2 mm。(g-l)测定了NIL-12植株第4幼节AR1和AR2的平均长度、横截面积、皮层面积、中柱面积或木质部/中柱比。AR-不定根;NIL-近等基因系。
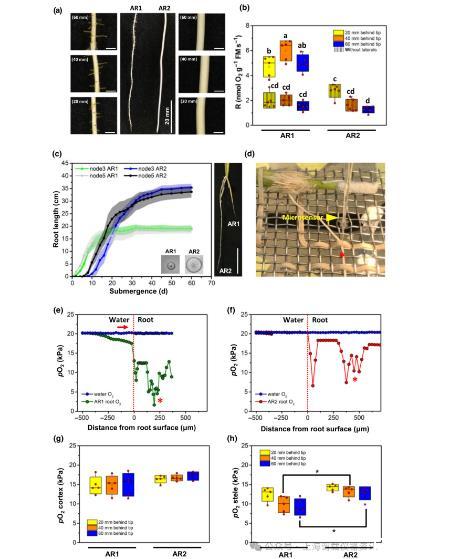
图4、水稻AR1和AR2供氧量和呼吸速率的比较(a)AR1发育侧根,AR2不发育侧根。(b)在距离根尖不同距离处测定未(去除侧根)或有侧根(原始状态)的AR1和未在这些位置形成侧根的AR2的呼吸速率。从浸入水中的NIL-12植物的8-10厘米长的根中收集根尖(50-70毫米)处20毫米(10-30毫米)、40毫米(30-50毫米)和60毫米的根段。对不同根类型和位置的呼吸速率进行对数转换数据的单因素方差分析和Tukey检验AR1在所有位置n=5,AR2在距离根尖20和40 mm,位置n=6,距离根尖60 mm位置n=5。(c)连续测定第3和第5年轻节点(节点3和5)的AR1和AR2的平均长度。淹没60天后,AR1和AR2在NIL-12的节点5生长。(d)一个固定的AR浸泡在水中,用距离尖端60毫米处的微传感器测量氧气(红星)。AR连接的杆的切割端用硅胶管和小玻璃珠密封。为了使氧气能够通过阀杆扩散供应,硅管被刺穿,并与气罐上方的空气接触,同时防止坑腔被淹没。大量的水被空气轻轻净化,以保持氧张力接近大气平衡,并维持稳定的扩散边界层。(e,f)距离AR1(e)或AR2(f)尖端60mm处的径向氧分布。红色箭头表示氧气从水中扩散到根部。红色星号表示中央石碑。(g,h)中位皮层(g)或中柱(h)pO2,由每种根类型的n=5个径向O2剖面计算,描绘为盒状须状图,显示平均值(+)、中位值(水平线)、50%四分位数(盒)、最小值和最大值(柱),圆圈表示单个数据点。根类型和位置之间的统计比较采用双向方差分析Tukey测试显示,AR1和AR2在距离尖端40和60 mm处的中柱O2有显著差异。AR-不定根;NIL-近等基因系。
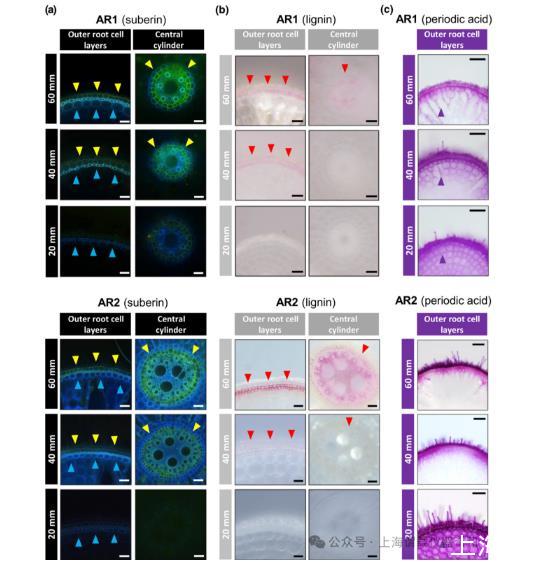
图5、在水稻中,AR2构成了气体扩散屏障,而AR1不构成气体扩散屏障。(a)在距离根尖的距离处显示AR1或AR2的木栓体染色。由黄色箭头所示的黄-绿颜色可见垫层片层。蓝色箭头指向细胞壁自动荧光。(b)AR1或AR2的木质素染色可见粉红色,并用红色箭头表示。棒材,20μm。(c)在AR1中酸通过外质体发生周期性扩散,但在AR2中没有。紫色箭头表示检测到周期性酸的皮层细胞层。棒材,50μm。AR=不定根。
结论与展望
本论文报道了两种类型的水生不定根的形成,在出现的时间和关键的形态学和解剖学特征上有显著差异。相关研究发现,叶鞘延迟了淹水后根系发育相关基因的时空表达,决定了根系出现的时间,甚至在淹水后期提高了根系的粗细和伸长能力。研究人员为了深水水稻的不定生根部的氧浓度分布,使用O2微传感器测量附着在水稻茎节上的水生不定根的径向O2分布。研究表明深水水稻中的AR2可以作为一种进化防御策略来应对周期性和长期的淹没。形态和解剖特征表明,AR2比AR1更适应水淹。叶鞘是影响AR2生长速率、伸长能力和根径的关键因素。与水稻(T65)相比,NIL-1和NIL-12在淹水过程中产生更多的AR2,说明控制AR2发育的关键基因包含在QTL1和QTL12中。研究人员展望下一步研究这两个qtl中调控AR2生长的基因。此外AR2在部分淹水过程中关于水和养分吸收的功能必须被阐明,以证明在经历长期浅淹的地区引入水生不定根的现代品种是合理的。
 相关新闻
相关新闻