热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
神经细胞电生理技术主要包括:膜片钳技术(patch clamp recording technique,PCRT)和在体多通道微电极阵列神经信号技术(In vivo multi-channel microelectrode array for neural signal recording technique,M-NEMEA)。
在体多通道同步记录技术是采用细胞外记录的方法来检测神经元群的同步电活动。该系统包括微电极阵列、数据采集和分析系统。应用这一技术可同步记录多个脑区的大量神经元的电活动,研究不同脑区的神经元放电在时间和空间上的联系,进而通过分析神经元的放电模式来研究大脑对外部事件的编码机制〔20〕。
多通道在体记录技术,能在自由活动的动物脑内,观察和记录大脑局部区域几十个、上百乃至上千个神经元的活动状况,是目前作为分析群体神经元在网络水平编码机制的有效工具〔21〕。John C Lilly在1949年采用多电极阵列植入的方法研究灵长类的行为和神经元的放电关系,利用了25个微电极记录到了610个神经元的放电〔22〕。在20世纪中期,研究人员用毛细玻璃管拉制了玻璃微电极。玻璃微电极的发明,使研究者可以将电极尖端插入神经细胞内,记录细胞体或轴突、树突内的电位变化,推动了神经电生理学细胞水平的研究。
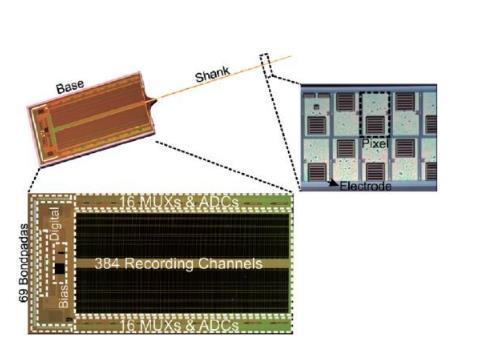
体多通道微电极阵列神经信号技术的技术特点
M-NEMEA通过离体神经网络活动的多通道电生理同步检测的方法,可研究神经网络群体特征的电生理特性,是目前研究神经网络发育和功能特征的重要方法。随着计算机技术的发展和微加工技术的提高,微电极阵列技术已经广泛应用于离体培养神经网络放电模式的检测中,从而为研究神经突触的可塑性,神经网络功能连接的形成与再生,以及神经网络发育的规律及生理节律提供了技术支持和保障。目前用于体外神经网络电生理检测的微电极阵列及多通道电生理检测系统多购自国外厂家,具有软件操作界面人性化,系统做工精良,系统稳定,处理功能强大等优点,但也存在不足。例如购买价格昂贵;易损耗,微电极阵列(microelectrode array,MEA)重复利用率低,多次使用后,表面电极材料会被损耗,影响信号记录;商业化系统的功能相对固定,不易进一步升级,不能完全满足实验室实验需求〔24〕。
M-NEMEA在视神经细胞的研究及应用
M-NEMEA作为细胞外记录的方法,通过对神经元群同步电活动的检测,可以同时采集大量细胞的动作电位的详细信息,从而为研究大脑神经细胞及细胞之间相互作用提供了重要的技术支持〔23〕,具有重要的意义。Tusa等〔25〕人利用微电极细胞外记录的方法对猫初级视皮层感受野进行系统的绘测,绘制出皮层17区和视野的拓扑对应关系图。陈昕等〔26〕采用玻璃微电极在猫的初级视皮层17区进行细胞外记录,并结合光学成像法定位猫初级视皮层17区不同部位的视野拓扑的离心度,结果表明神经元的感受野的离心度与光学记录到的结果十分接近,从而为大面积确定皮层细胞感受野在视野中的位置提供了快速和较准确的方法。史学锋等〔27〕采用微电极阵列神经细胞信号采集技术对斜视性弱视猫行细胞电生理检测,观察视皮层21a区神经细胞眼优势的相关变化,结果发现与正常组相比,模型组21a双眼细胞比例明显下降,在纹外视皮层斜视性弱视的病理不足更为明显,神经细胞眼优势明显转移至正常眼。姚军平等〔28〕用微电极阵列神经细胞信号采集技术检测22 d正常大鼠(Long Evans大鼠,下同)、22 d单眼剥夺大鼠、100 d正常大鼠、100 d单眼剥夺大鼠眼优势柱的分布,结果发现22 d正常大鼠、100 d正常大鼠、100 d单眼剥夺大鼠视皮层眼优势柱分布为对侧眼优势,22 d单眼剥夺大鼠视皮层眼优势柱分布为同侧眼优势,表明正常视觉环境下发育的Long Evans大鼠,眼优势柱分布为对侧眼优势,其在幼年期视皮层具有可塑性,在异常的视觉环境下可影响其眼优势柱的分布;而在成年时,Long Evans大鼠视觉可塑性终止,异常的环境不能影响其眼优势柱的分布。刘虎等〔29〕用微电极阵列神经细胞信号采集技术检测斜视性弱视猫纹状皮层细胞功能和放电水平,结果发现较正常组相比,模型组皮层细胞数量明显减少,眼优势未转移;模型组与正常眼相比眼驱使皮层细胞的高端截止空间和最优势空间频率显著下降,表明驱使皮层细胞的空间特性和两眼拮抗作用是导致斜视性弱视的原因之一。谢芳等〔30〕用微电极阵列神经细胞信号采集技术记录幼猫初级视皮层V1区峰电位信号和场电位信号,观察眼优势指数的变化,结果发现场电位和锋电位的眼优势指数呈正相关,表明双眼视刺激的平衡程度直接影响神经元的整合作用。徐鹏景等〔31〕通过在体胞外记录的方法研究视觉系统中二阶信号和一阶信号是由相同还是不同的通路来完成的。发现大多数膝外体细胞对二阶信号存在调制反应,但反应强度低于一阶信号,并且二阶信号反应中的非线性成分与线性成分的比值高于一阶信号反应,表明猫外膝体上一阶信号和二阶信号的处理可能是由两类不同的传递通路来完成的。
丁娟等〔32〕将健康Wistar大鼠48只分2组进行不同目的研究。第一组健康Wistar大鼠24只分为4组,每组6只,除正常组外,其余各组在出生后14(P14)天制作右眼单眼形觉(MD)模型。采用细胞外记录技术观察各组视皮质II/III层突触传递LTP诱发率,以及各组在高频刺激(HFS)刺激后,兴奋性突触后电位(EPSP)及群峰电位的变化。结果表明,①各视皮质II/III层LTP诱发率正常P45天组最高(5/6),最低的为MDP90天组。②高频刺激后EPSP斜率和群峰电位(PS)幅值不同时段与基础突触传递水平的比较:EPSP斜率和PS幅值在正常P45天组、正常P90天组HFS后0、30、60 min提高显著(P<0.01)。同一时间点比较,HFS后30、60 min各组EPSP斜率以及PS幅值与基础值之比,组间比较具有显著性差异(P<0.01)。从而表明大鼠皮质LTP诱发呈视觉依赖性,MD可降低剥夺眼对侧视皮质LTP诱发率;EPSP斜率,PS幅值对高频刺激的反应受年龄和异常视觉经验影响,年龄越大,以及视觉经验的异常(单眼形觉剥夺)会降低视皮质对高频刺激的反应。第二组仍采用细胞外记录技术观察各组注药后10、30、60、90 min对基础突触传递水平的影响。给药90 min后,给予视皮层质Ⅳ层高频刺激,观察药物对视皮质Ⅱ/Ⅲ层LTP诱导的影响。结果表明:①不同浓度天然药物A对基础突触传递水平的影响:在MD对照组和正常组侧脑室给予人工脑脊液(ACSF)后10、30、60、90 min EPSP斜率和PS幅值并无明显变化,天然药物A随给药时间延长能提高EPSP斜率和幅值,高剂量组更具有显著性。②天然药物A对高频刺激诱导的Ⅱ/Ⅲ层LTP的影响:HFS 30、60 min后,EPSP斜率和PS幅值MD高、低剂量给药组、正常组高于MD对照组(P<0.01)。从而表明:天然药物A侧脑室注药能提高MD成年大鼠视皮质Ⅱ/Ⅲ层基础突触传递水平,增强EPSP以及PS幅值,呈剂量依赖性加强HFS对视皮质Ⅱ/ⅢLTP的诱导,提高突触传递效能。
