热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
研究简介:Methanococcus maripaludis是一种嗜温产甲烷菌,能够利用氢气(H₂)或甲酸盐(formate)将二氧化碳(CO₂)还原为甲烷(CH₄)以获取能量。这种菌在盐沼沉积物中很常见,具有多种氢化酶,这些酶在氢气代谢中起重要作用。由于氢气是一种清洁且高效的能源,研究人员希望探索这种产甲烷菌利用甲酸盐产生氢气的潜力,以期为生物制氢提供新的思路。以甲酸盐或H2生长的静息细胞的比活性为0.4至1.4 U·mg-1(干重)。甲酸盐生长的细胞中H2生产遵循米氏动力学,甲酸盐的半最大活性浓度(Kf)为3.6 mM。相比之下,H2生长的细胞中此过程遵循S型动力学,Kf为9 mM。甲酸盐依赖的H2生产的关键酶是甲酸脱氢酶Fdh。在缺失编码Fdh1同工酶的基因的突变体中,H2生产和生长严重减少,表明Fdh1是主要的甲酸脱氢酶。相反缺失编码Fdh2同工酶的基因的突变体具有接近野生型的活性,表明该同工酶不起主要作用。缺失辅酶F420还原氢化酶Fru的突变体的H2生产也严重减少,表明主要的H2生产途径由Fdh1和Fru组成。由于Δfru-Δfrc突变体保留了10%的野生型活性,因此还存在另一条途径。缺失编码F420依赖的亚甲基-H4 MTP脱氢酶(Mtd)或H2形成的亚甲基-H4 MTP脱氢酶(Hmd)的突变体活性也降低,表明第二条途径由Fdh1-Mtd-Hmd组成。与H2生产不同,这些突变体中甲烷生成的细胞速率未受影响,表明观察到的H2生产并非甲烷生成的直接中间产物。甲酸盐依赖的高速率H2生产展示了M.maripaludis从甲酸盐微生物生产H2的潜力。
Unisense氢气微电极系统的应用
Unisense微电极被用于精确测量Methanococcus maripaludis在厌氧条件下从甲酸盐产生氢气(H₂)的浓度和速率。将细胞悬浮液加入到含有Unisense微电极的反应比色皿中,通过改良的安培式氧气克拉克型电极连接到Unisense picoammeter PA2000,实时监测溶解氢气的浓度。通过对Unisense微电极测量数据分析发现,M.maripaludis在最初的1分钟内从甲酸盐产生氢气的速率呈线性增长,随后速率下降直至达到平衡。氢气产生的最大浓度在0.4到0.8 mM之间,接近理论平衡浓度3 mM。
实验结论
M.maripaludis能够高效地从甲酸盐产生氢气,其产氢速率在不同生长条件下表现出显著差异。在最佳条件下,产氢速率可达1.4 U/mg(干重),这在已知的微生物中属于较高水平。氢气产生的速率和动力学特性受到生长条件的显著影响,尤其是氢气的可用性。在氢气限制条件下生长的细胞表现出更高的产氢活性。通过基因敲除实验,研究发现Fdh1是甲酸盐依赖性氢气产生的主要酶,而Fdh2的作用较小。Fdh1缺失的突变株在甲酸盐利用和氢气产生方面表现出严重缺陷,而Fdh2缺失的突变株则接近野生型水平。甲酸盐依赖的氢气产生遵循米氏动力学(Michaelis-Menten kinetics),其半饱和常数(Kf)为3.6 mM,表明Fdh1对甲酸盐的亲和力较高。氢气产生的速率与甲烷生成的速率相当,但实验结果表明,氢气并非甲烷生成过程中的必需中间体。在某些突变株中,氢气产生显著减少,但甲烷生成并未受到显著影响。这表明M.maripaludis在甲酸盐代谢中可能存在其他电子传递途径,氢气的产生可能是代谢过程中的副产物。
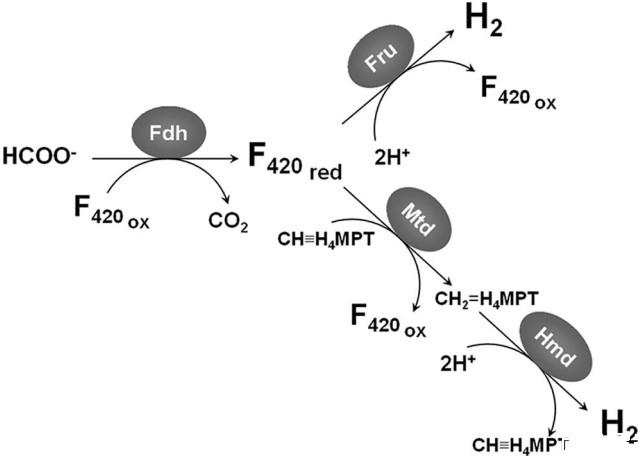
图1、氢营养型产甲烷菌中H2合成的潜在途径。第一条途径包含Fdh和Fru。在该途径中,Fdh还原F420。在含硒培养的细胞中,F420H2被[Ni-Fe]氢化酶Fru氧化。第二条途径中,F420H2被Mtd氧化,将亚甲基-H4MPT还原为亚甲基-H4MPT,随后被不含镍的氢化酶Hmd重新氧化产生H2。
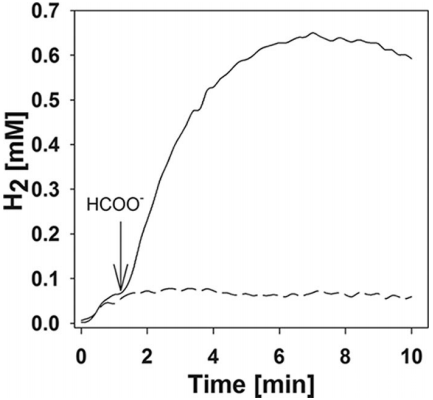
图2、M.maripaludis静息细胞从甲酸盐产生H2的初始速率(实线)。细胞以甲酸盐培养,经缓冲液洗涤后重悬于反应比色皿中。通过添加20 mM甲酸钠(箭头)启动反应。虚线显示未添加甲酸盐的细胞悬浮液孵育结果。
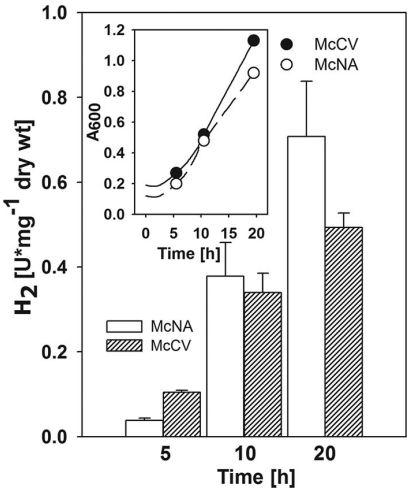
图3、H2限制条件下生长的静息细胞甲酸盐依赖H2生产的变化。野生型S2在1升Wheaton瓶中用100 ml肉汤培养,分别使用基本培养基McNA(○)和丰富培养基McCV(●)。实验开始时,瓶子用138 kPa H2-CO2(80:20[体积比])加压。在不同吸光度时采集细胞样品(20 ml),如插图中生长曲线上的点所示。每次取样后,瓶子用N2-CO2(80:20[体积比])重新加压。这导致H2限制但维持CO2浓度。
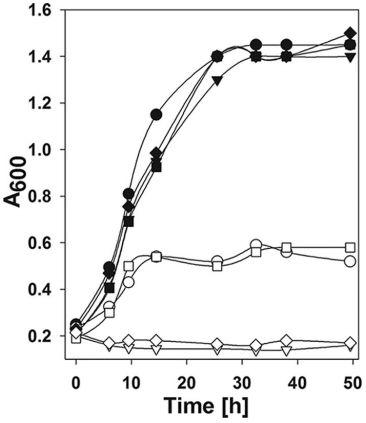
图4、野生型和fdh突变株在H2或甲酸盐条件下的生长情况。实心符号表示H2(276 kPa;H2-CO2 80:20[体积比])条件下的生长;空心符号表示甲酸盐(100 mM)条件下的生长。圆形代表S2;倒三角形代表ΔfdhA1;方形代表ΔfdhA2;菱形代表ΔfdhA1-ΔfdhA2。每个数据点代表两次重复实验的平均值。
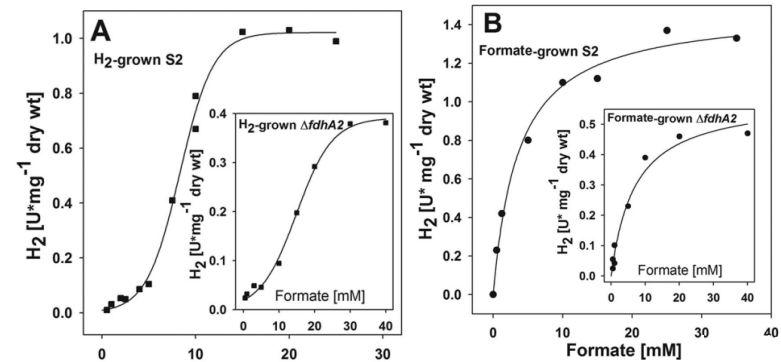
图5、野生型菌株S2静息细胞的甲酸盐依赖性H2生产动力学,该菌株先前分别在H2(A)或甲酸盐(B)条件下培养。ΔfdhA2突变体的动力学如插图所示。S2和ΔfdhA2突变株分别培养至吸光度约0.8和0.6。
结论与展望
本论文研究了嗜温产甲烷菌Methanococcus maripaludis利用甲酸盐(formate)产生氢气(H₂)的特性及其潜在应用。M.maripaludis是一种能够利用氢气或甲酸盐将二氧化碳还原为甲烷的产甲烷菌,其在盐沼沉积物中广泛存在,并且具有多种氢化酶,这些酶在氢气代谢中起重要作用。本研究旨在探索这种菌利用甲酸盐产生氢气的能力,为生物制氢提供新的思路。研究人员使用了M.maripaludis的野生型菌株S2及其突变株。菌株在含有甲酸盐或氢气的培养基中生长,实验中还使用了不同的培养基成分和气体压力条件。通过定制的厌氧水浴比色皿和改良的安培式氧气克拉克电极(unisense)测量溶解氢气的浓度,同时通过气相色谱法检测甲烷的产生。此外研究还制备了细胞提取物,并通过光谱法测定甲酸脱氢酶(Fdh)和F420还原氢化酶的活性。Unisense微电极在本研究中被用于精确测量Methanococcus maripaludis在厌氧条件下从甲酸盐产生氢气(H₂)的浓度和速率。通过分析unisense精确测量氢气的产生速率和浓度,为研究M.maripaludis的氢气代谢机制提供了关键数据,。本论文深入探讨了M.maripaludis在生物制氢方面的潜力,还揭示了其氢气产生机制,为未来开发高效的生物制氢系统提供了重要的理论基础和实验数据。
