热线:021-66110810,66110819
手机:13564362870
热线:021-66110810,66110819
手机:13564362870
微电极测定的H2的来源很可能是输送结肠发酵产生的H2的血管系统,考虑到小鼠肝脏(通过手术)部分暴露于空气中,测量值的巨大差异也就不足为奇了。考虑到小鼠肝脏(通过手术)部分暴露于空气中,测量值的巨大差异也就不足为奇了。啮齿动物肠道排出氢气的情况已有记载,但我们发现的与活体小鼠特定组织(胃和肝脏)相关的气体水平是意料之外的。当然,这种体积小、易扩散但能量高的底物对肝吸虫肝脏定殖是否有用,必须等待使用缺乏H2利用能力的肝吸虫基因突变体进行关键动物研究后才能确定。
表1记录了肝吸虫的H2-O2呼吸途径仅为幽门螺杆菌的约10%(基于每个细胞)。在厌氧条件下,由于没有观察到H2的吸收(或释放),因此整个细胞的H2氧化取决于氧气的加入。
H.hepaticus全细胞能够将H2氧化偶联到O2呼吸消耗。厌氧条件下未观察到H2吸收。我们立即注意到H2氧化的速率低于我们对幽门螺杆菌的常规观察。为了严格解决这一观察结果,将肝吸虫的H2氧化活性与在相同条件下生长的两种常见幽门螺杆菌菌株(在含10%H2的气氛中培养的血液琼脂平板)的H2氧化能力进行了比较。据报道,摄取型氢化酶经历厌氧活化(有时称为还原活化)现象。其特征是当条件是高度还原或厌氧时,酶活性高得多。当O2存在于反应混合物中时,H.hepaticus酶的厌氧H2氧化率仅为16%(表1)。这种厌氧激活量类似于我们对幽门螺杆菌细胞的观察(表1)和我们之前对从临床幽门螺杆菌分离物获得的膜的观察。对于已经详细研究了这种厌氧活化的系统,在还原条件下实现完全活性与电子受体还原的速率(即氢化酶周转率)有关。
表二,氢化酶的电子受体特异性。
幽门螺杆菌(膜)和肝吸虫(渗透细胞)的100%活性(亚甲蓝率)分别为382和62nmol/min/mg蛋白质。
如表1脚注a所述制备肝吸虫透化细胞,并在加入1%十二烷基硫酸钠后,用双辛可宁酸(Pierce Chemical Co)测量蛋白质,然后加热细胞(90°C,10分钟)。在50M连二亚硫酸钠存在下,在饱和水平厌氧测定所有电子受体活性。
肝吸虫中氢化酶的特征与幽门螺杆菌的报道相似,因此肝吸虫系统很可能也是膜结合的,并参与节能。它可能涉及“H2分裂”步骤后其他膜相关电子传递蛋白的还原。因此,确定了该H2氧化系统发挥作用的染料介导的氧化还原电势范围。
通常,H2摄取氢化酶与正电位的氧化还原受体一起起作用,但与负氧化还原电位受体不起作用。至于幽门螺杆菌,H.hepaticus酶能够将H2氧化与正受体的还原偶联(表2),但不能将负电位受体苄基紫精偶联。这些实验是在没有氧气的情况下进行的。幽门螺杆菌和肝吸虫系统之间的一个有趣的区别是细胞色素c作为氢化酶受体的适用性。中等氧化还原电位的细胞色素c是肝吸虫氢化酶的良好受体;这可能意味着肝吸虫中来自氢化酶的电子的体内受体比幽门螺杆菌膜中的初始受体处于更高的氧化还原电位(醌或细胞色素)。我们之前已经表明,幽门螺杆菌中的氢氧化与细胞色素还原有关,这些含血红素的成分在末端(O2结合)氧化酶的还原(12)。然而,由于H2氧化膜,这些差异光谱实验对肝吸虫来说是不可能的无法获得颗粒。膜分离是以限制氧气的方式进行的(在细胞破坏期间和之后使用充氩缓冲液,以及通过充氩注射器转移提取物),但不是在严格的厌氧条件下进行的,因此在H2-O2呼吸链中可能存在O2不稳定因子。然而,由于H2氧化与O2吸收相耦合,并且与正电位染料的还原相耦合,因此在H.hepaticus膜中可能存在(尚未确定的)含有血红素的组分,这些组分参与保存最初由H2提供的电子的能量。
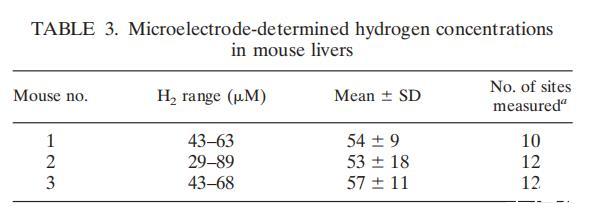
我们使用改良的微电极模型氢气-50(unisense),用微电极法测定了成年活体小鼠肝脏中的氢浓度(表1)。电极的尖端直径为50μM。雌性小鼠C57BL使用氟烷麻醉。在整个过程中,小鼠保持存活但完全麻醉,并在最后一次测量后立即进行安乐死。大多数测量是在探针进入肝脏不到1.0mm时进行的,以免损坏探针。实际上,在最初的一些测量中,当探针插入组织更深处时,探针已受到损坏。在信号稳定(约5至7秒)后记录氢气读数。每只小鼠肝脏最多可在12个部位重复上述步骤,使用校准室获得氢气的标准曲线。平均氢含量超过50μM,比表观Km高出20倍,并略高于报道的小鼠胃中的氢气含量。组织氢气测量结果共同支持了一个假设,即动物体内的许多组织中都存在氢气,因此有必要进一步考虑氢气作为能量库供宿主动物体内感染性细菌使用的可能性。
微电极测定的氢气来源很可能是血管系统输送的结肠发酵产生的氢气,考虑到部分小鼠肝脏(通过手术)暴露于空气中,测量值的巨大差异不足为奇。据推测,暴露在空气中测量到的水平会低于完整动物体内的水平。啮齿类动物肠道排出氢气的情况已有记载,但我们在活体小鼠的特定组织(胃和肝脏)中发现的气体水平是意料之外的。当然,这种体积小、易扩散但高能量的底物对肝门螺杆菌肝脏定殖的作用必须通过使用缺乏氢气利用能力的肝门螺杆菌基因靶向突变体进行关键的动物研究。
 相关新闻
相关新闻